

Thalamic miR-338-3p mediates auditory thalamocortical disruption and its late onset in models of 22q11.2 microdeletion
- PMID: 27892953
- PMCID: PMC5218899
- DOI: 10.1038/nm.4240
Thalamic miR-338-3p mediates auditory thalamocortical disruption and its late onset in models of 22q11.2 microdeletion
Abstract
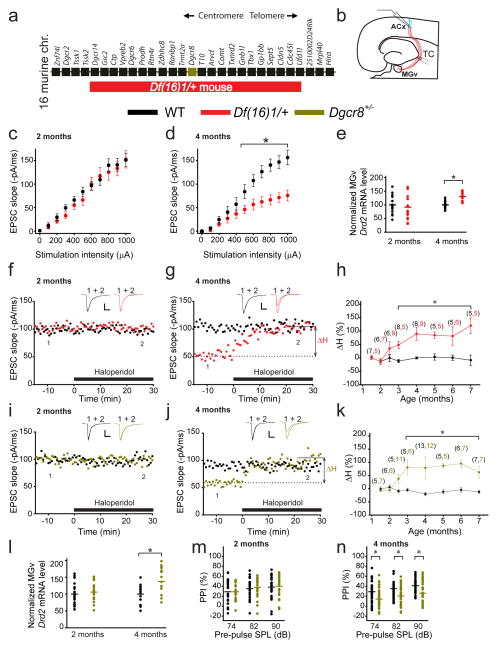
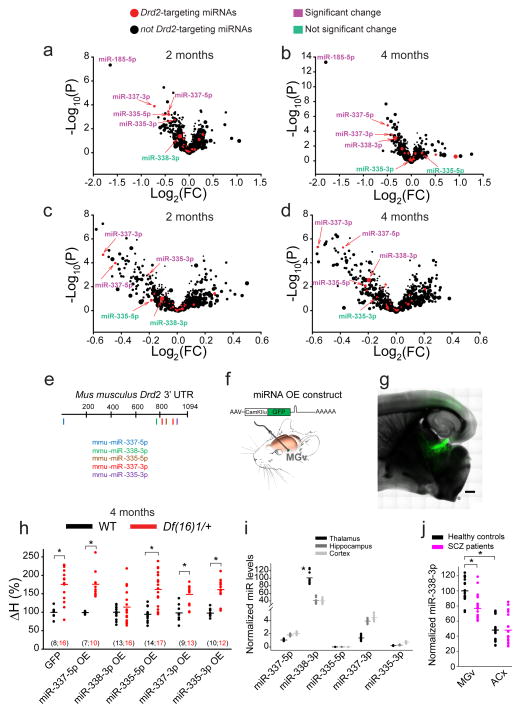
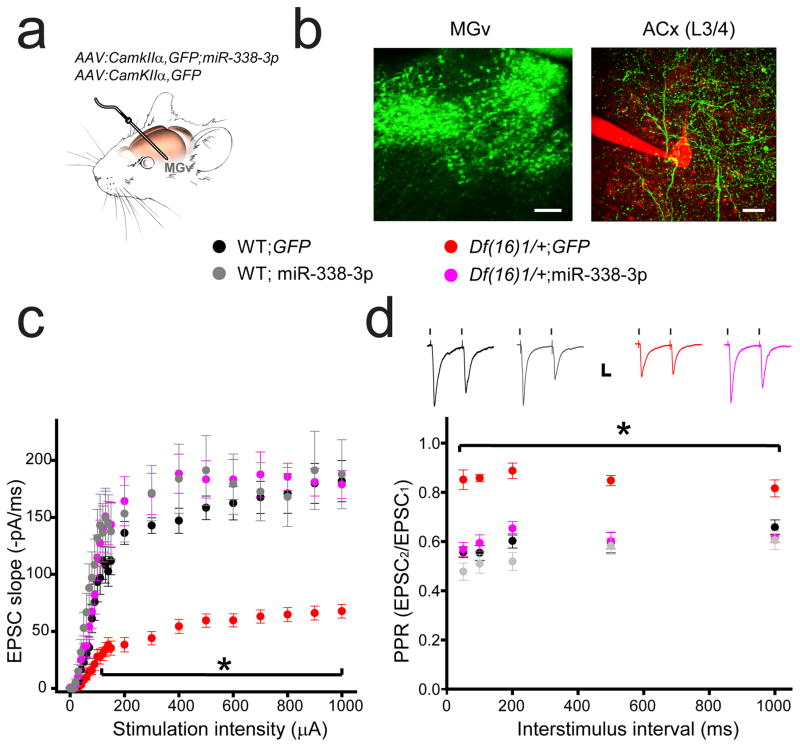
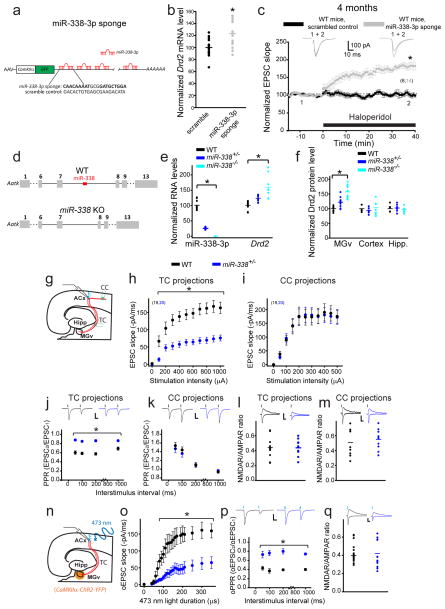
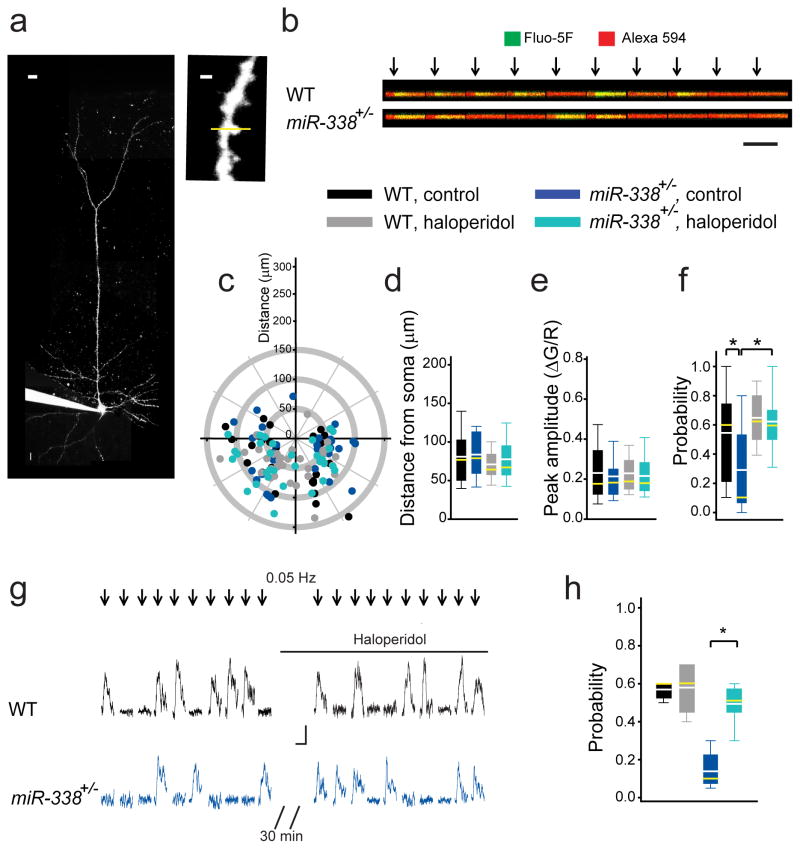
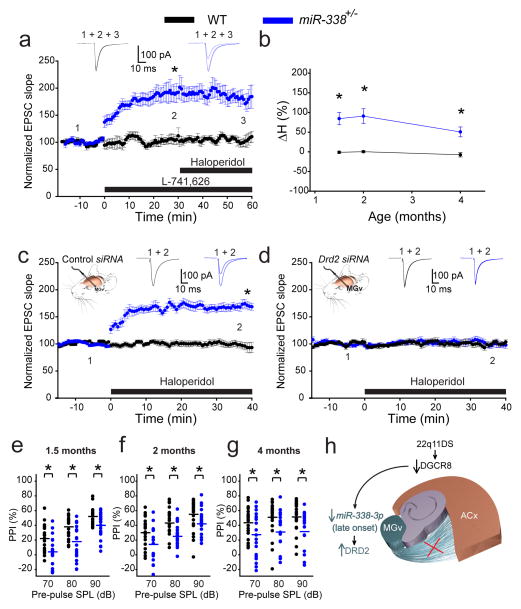
Although 22q11.2 deletion syndrome (22q11DS) is associated with early-life behavioral abnormalities, affected individuals are also at high risk for the development of schizophrenia symptoms, including psychosis, later in life. Auditory thalamocortical (TC) projections recently emerged as a neural circuit that is specifically disrupted in mouse models of 22q11DS (hereafter referred to as 22q11DS mice), in which haploinsufficiency of the microRNA (miRNA)-processing-factor-encoding gene Dgcr8 results in the elevation of the dopamine receptor Drd2 in the auditory thalamus, an abnormal sensitivity of thalamocortical projections to antipsychotics, and an abnormal acoustic-startle response. Here we show that these auditory TC phenotypes have a delayed onset in 22q11DS mice and are associated with an age-dependent reduction of miR-338-3p, a miRNA that targets Drd2 and is enriched in the thalamus of both humans and mice. Replenishing depleted miR-338-3p in mature 22q11DS mice rescued the TC abnormalities, and deletion of Mir338 (which encodes miR-338-3p) or reduction of miR-338-3p expression mimicked the TC and behavioral deficits and eliminated the age dependence of these deficits. Therefore, miR-338-3p depletion is necessary and sufficient to disrupt auditory TC signaling in 22q11DS mice, and it may mediate the pathogenic mechanism of 22q11DS-related psychosis and control its late onset.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
