

A High-Fat Diet Promotes Mammary Gland Myofibroblast Differentiation through MicroRNA 140 Downregulation
- PMID: 27895151
- PMCID: PMC5288574
- DOI: 10.1128/MCB.00461-16
A High-Fat Diet Promotes Mammary Gland Myofibroblast Differentiation through MicroRNA 140 Downregulation
Abstract
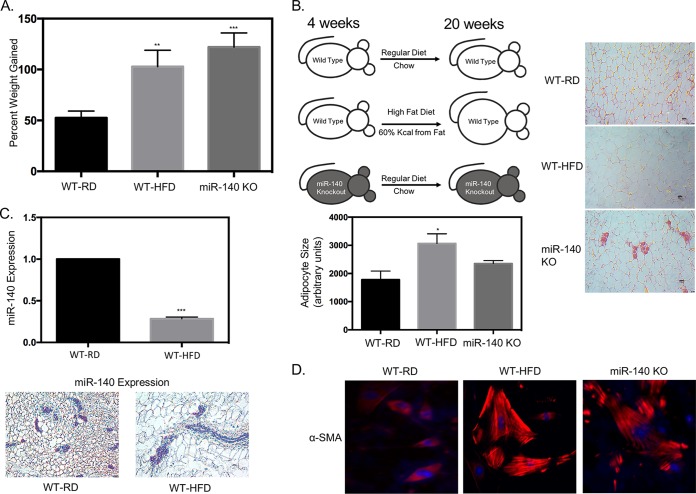
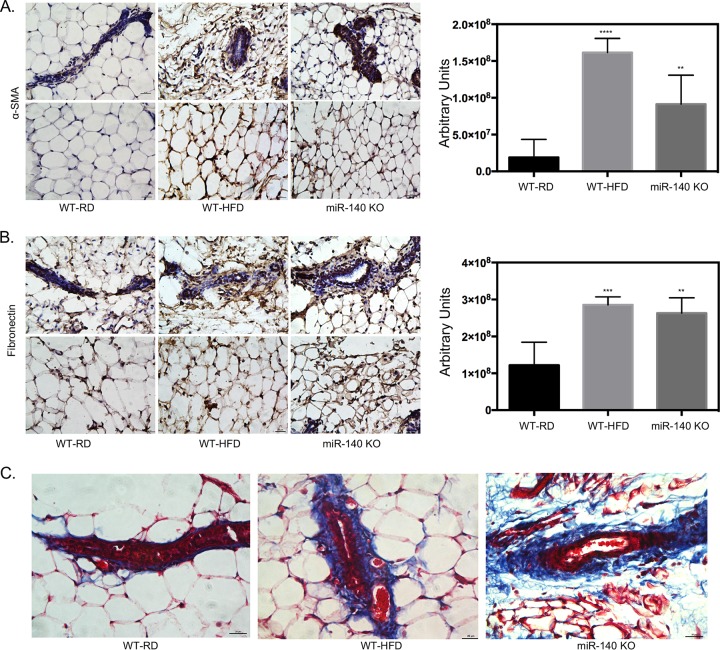
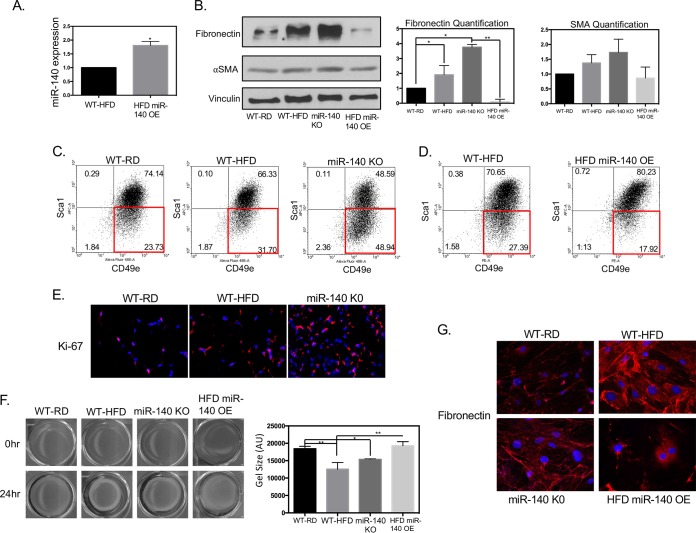
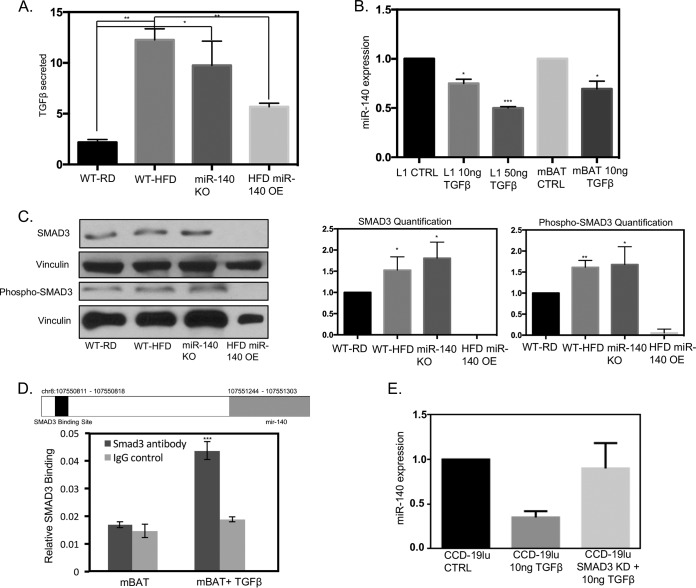
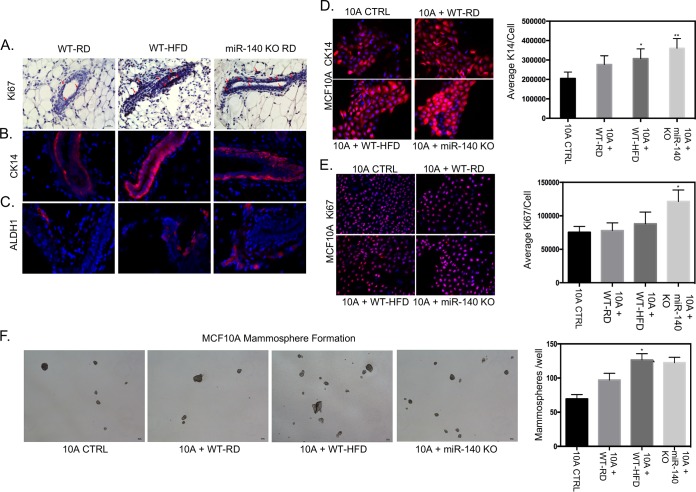
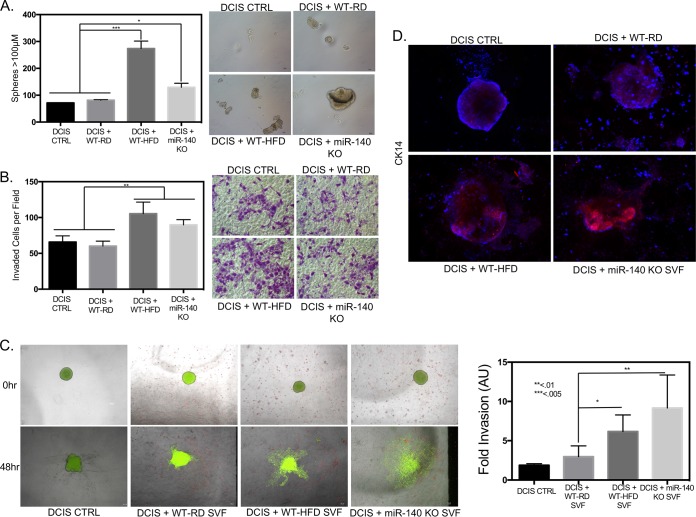
Human breast adipose tissue is a heterogeneous cell population consisting of mature white adipocytes, multipotent mesenchymal stem cells, committed progenitor cells, fibroblasts, endothelial cells, and immune cells. Dependent on external stimulation, adipose-derived stem cells differentiate along diverse lineages into adipocytes, chondrocytes, osteoblasts, fibroblasts, and myofibroblasts. It is currently not fully understood how a high-fat diet reprograms adipose-derived stem cells into myofibroblasts. In our study, we used mouse models of a regular diet and of high-fat-diet-induced obesity to investigate the role of dietary fat on myofibroblast differentiation in the mammary stromal microenvironment. We found that a high-fat diet promotes myofibroblast differentiation by decreasing microRNA 140 (miR-140) expression in mammary adipose tissue through a novel negative-feedback loop. Increased transforming growth factor β1 (TGF-β1) in mammary adipose tissue in obese mice activates SMAD3 signaling, causing phospho-SMAD3 to bind to the miR-140 locus and inhibit miR-140 transcription. This prevents miR-140 from targeting SMAD3 for degradation, resulting in amplified TGF-β1/SMAD3 signaling and miR-140 downregulation-dependent myofibroblast differentiation. Using tissue and coculture models, we found that myofibroblasts and the fibrotic microenvironment created by myofibroblasts impact the stemness and proliferation of normal ductal epithelial cells and early-stage breast cancer invasion and stemness.
Keywords: Obesity; breast cancer; fibrosis; miRNA; myofibroblasts.
Copyright © 2017 American Society for Microbiology.
Figures
References
-
- Chandler EM, Seo BR, Califano JP, Andresen Eguiluz RC, Lee JS, Yoon CJ, Tims DT, Wang JX, Cheng L, Mohanan S, Buckley MR, Cohen I, Nikitin AY, Williams RM, Gourdon D, Reinhart-King CA, Fischbach C. 2012. Implanted adipose progenitor cells as physicochemical regulators of breast cancer. Proc Natl Acad Sci U S A 109:9786–9791. doi: 10.1073/pnas.1121160109. - DOI - PMC - PubMed
-
- Kojima Y, Acar A, Eaton EN, Mellody KT, Scheel C, Ben-Porath I, Onder TT, Wang ZC, Richardson AL, Weinberg RA, Orimo A. 2010. Autocrine TGF-beta and stromal cell-derived factor-1 (SDF-1) signaling drives the evolution of tumor-promoting mammary stromal myofibroblasts. Proc Natl Acad Sci U S A 107:20009–20014. doi: 10.1073/pnas.1013805107. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials