

Reciprocal Modulation of IK1-INa Extends Excitability in Cardiac Ventricular Cells
- PMID: 27895596
- PMCID: PMC5108932
- DOI: 10.3389/fphys.2016.00542
Reciprocal Modulation of IK1-INa Extends Excitability in Cardiac Ventricular Cells
Abstract
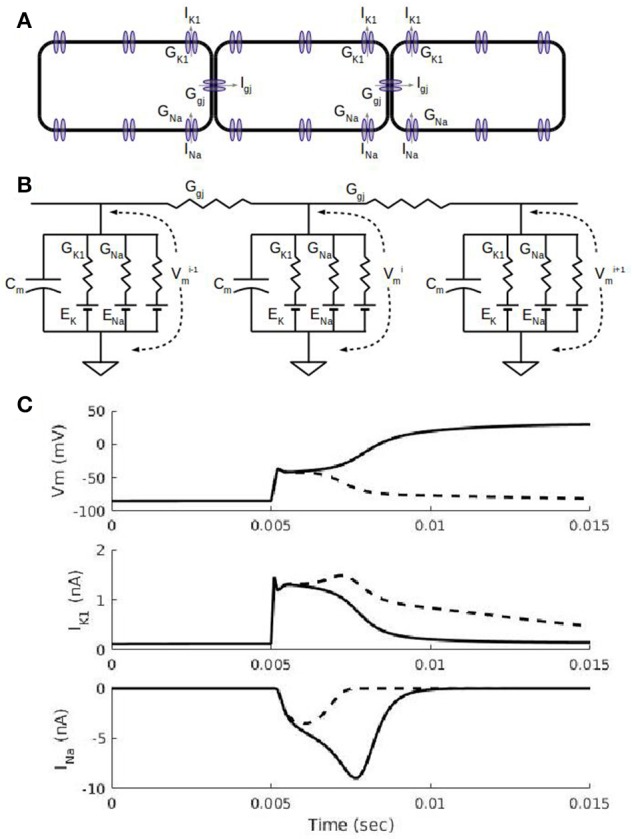
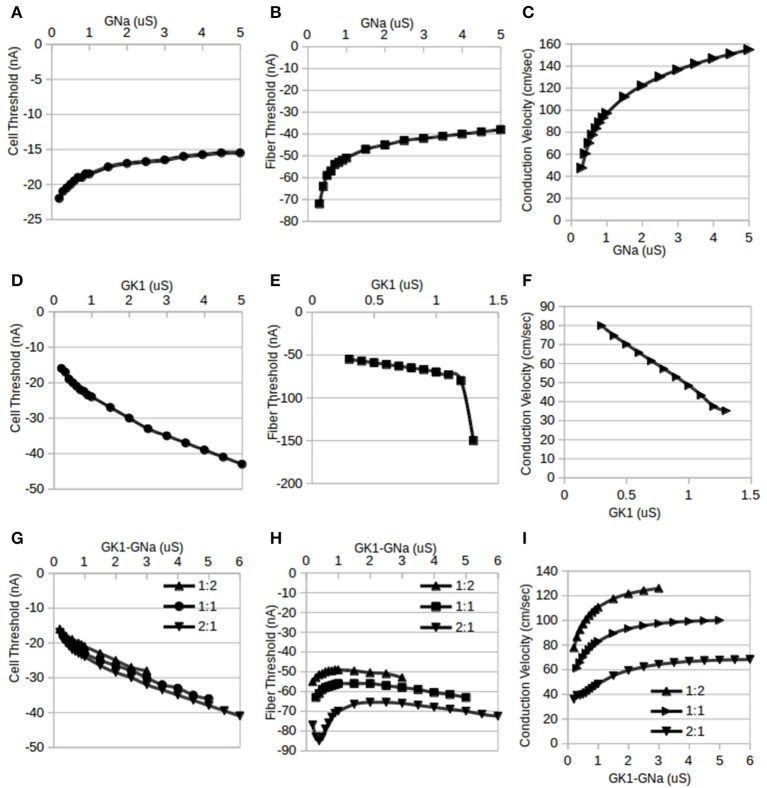
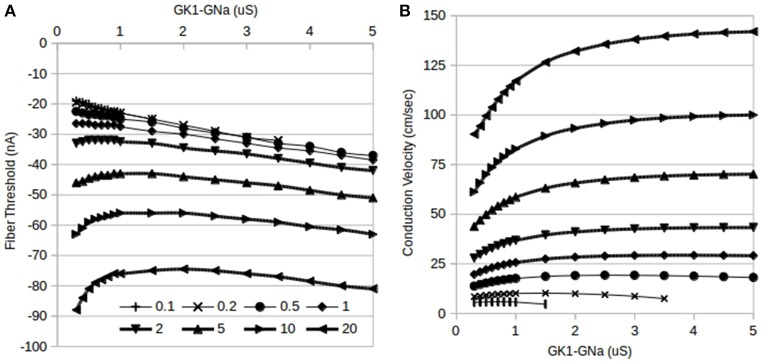
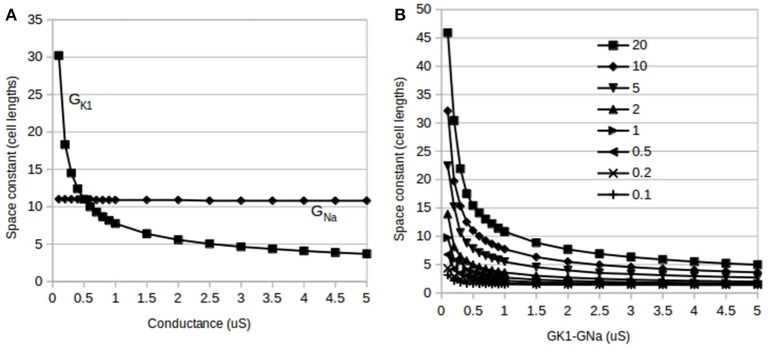
The inwardly rectifying potassium current (IK1) and the fast inward sodium current (INa) are reciprocally modulated in mammalian ventricular myocytes. An increase in the expression of channels responsible for one of these two currents results in a corresponding increase in expression of the other. These currents are critical in the propagation of action potentials (AP) during the normal functioning of the heart. This study identifies a physiological role for IK1-INa reciprocal modulation in ventricular fiber activation thresholds and conduction. Simulations of action potentials in single cells and propagating APs in cardiac fibers were carried out using an existing model of electrical activity in cardiac ventricular myocytes. The conductances, GK1, of the inwardly rectifying potassium current, and GNa, of the fast inward sodium current were modified independently and in tandem to simulate reciprocal modulation. In single cells, independent modulation of GK1 alone resulted in changes in activation thresholds that were qualitatively similar to those for reciprocal GK1-GNa modulation and unlike those due to independent modulation of GNa alone, indicating that GK1 determines the cellular activation threshold. On the other hand, the variations in conduction velocity in cardiac cell fibers were similar for independent GNa modulation and for tandem changes in GK1-GNa, suggesting that GNa is primarily responsible for setting tissue AP conduction velocity. Conduction velocity dependence on GK1-GNa is significantly affected by the intercellular gap junction conductance. While the effects on the passive fiber space constant due to changes in both GK1 and the intercellular gap junction conductance, Ggj, were in line with linear cable theory predictions, both conductances had surprisingly large effects on fiber activation thresholds. Independent modulation of GK1 rendered cardiac fibers inexcitable at higher levels of GK1 whereas tandem GK1-GNa changes allowed fibers to remain excitable at high GK1 values. Reciprocal modulation of the inwardly rectifying potassium current and the fast inward sodium current may have a functional role in allowing cardiac tissue to remain excitable when IK1 is upregulated.
Keywords: cardiac cells; mathematical model; reciprocal modulation.
Figures
Comment in
-
Commentary: Reciprocal Modulation of IK1-INa Extends Excitability in Cardiac Ventricular Cells.Front Physiol. 2016 Dec 23;7:647. doi: 10.3389/fphys.2016.00647. eCollection 2016. Front Physiol. 2016. PMID: 28066270 Free PMC article. No abstract available.
References
-
- Crawford J. D., Golubitsky M., Gomes M. G. M., Knobloch E., Stewart I. N. (1991). Boundary conditions as symmetry constraints, in Singularity Theory and its Applications: Warwick 1989, Part II: Singularities, Bifurcations and Dynamics, ed Roberts M. (Berlin: Springer; ), 63–79.
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous