

ALA-Induced Flavonols Accumulation in Guard Cells Is Involved in Scavenging H2O2 and Inhibiting Stomatal Closure in Arabidopsis Cotyledons
- PMID: 27895660
- PMCID: PMC5108921
- DOI: 10.3389/fpls.2016.01713
ALA-Induced Flavonols Accumulation in Guard Cells Is Involved in Scavenging H2O2 and Inhibiting Stomatal Closure in Arabidopsis Cotyledons
Abstract
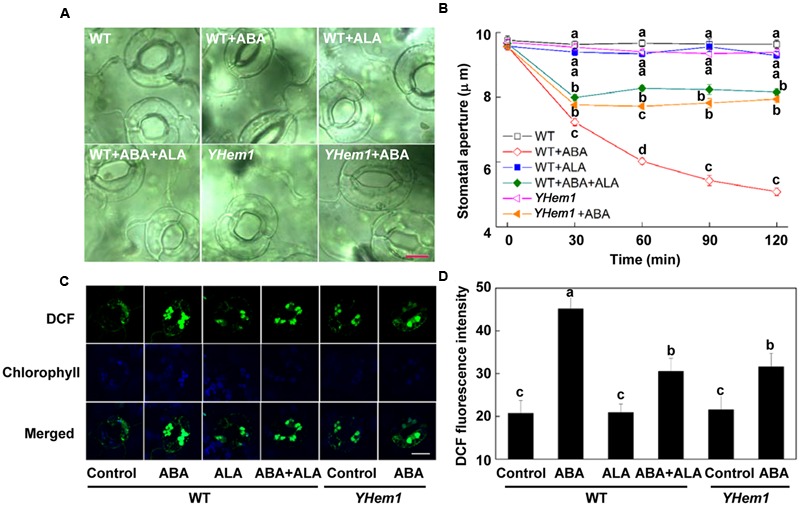
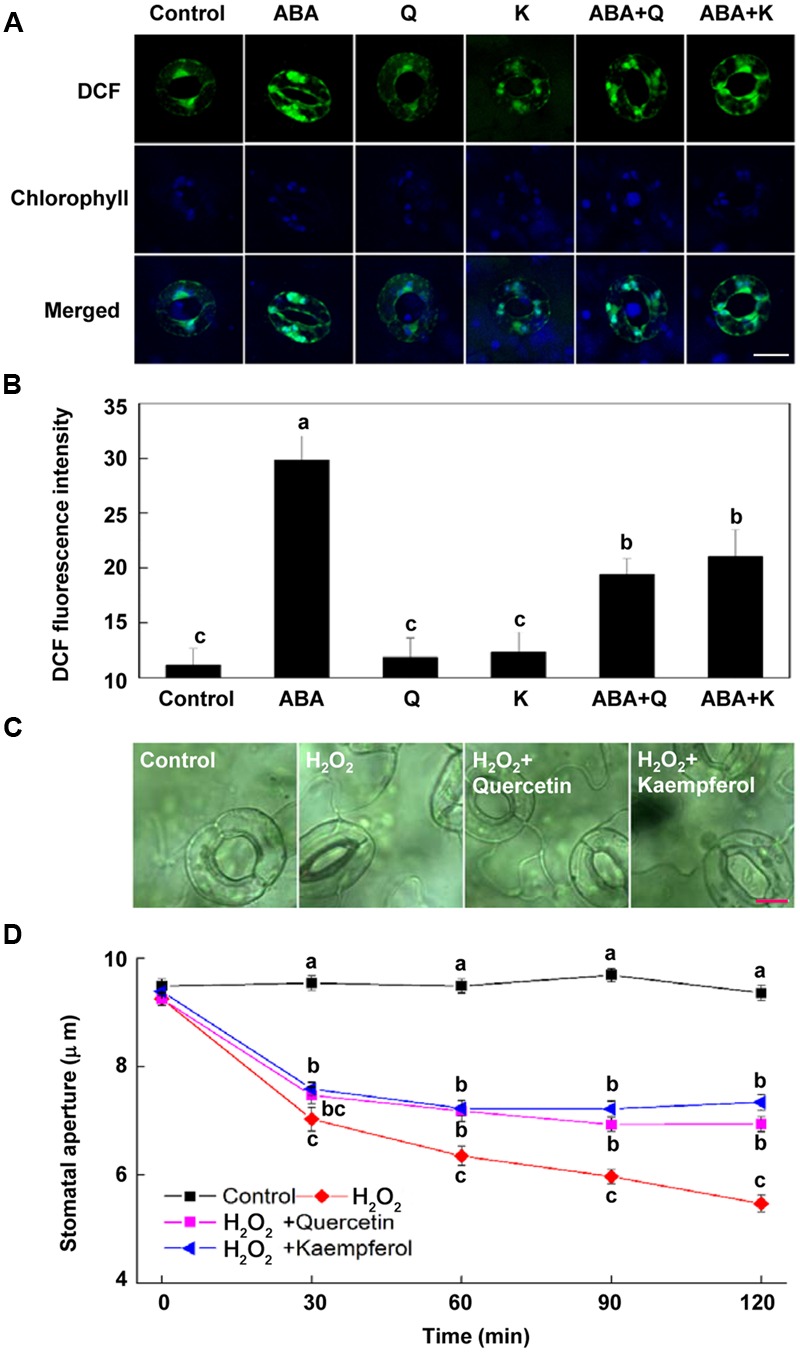
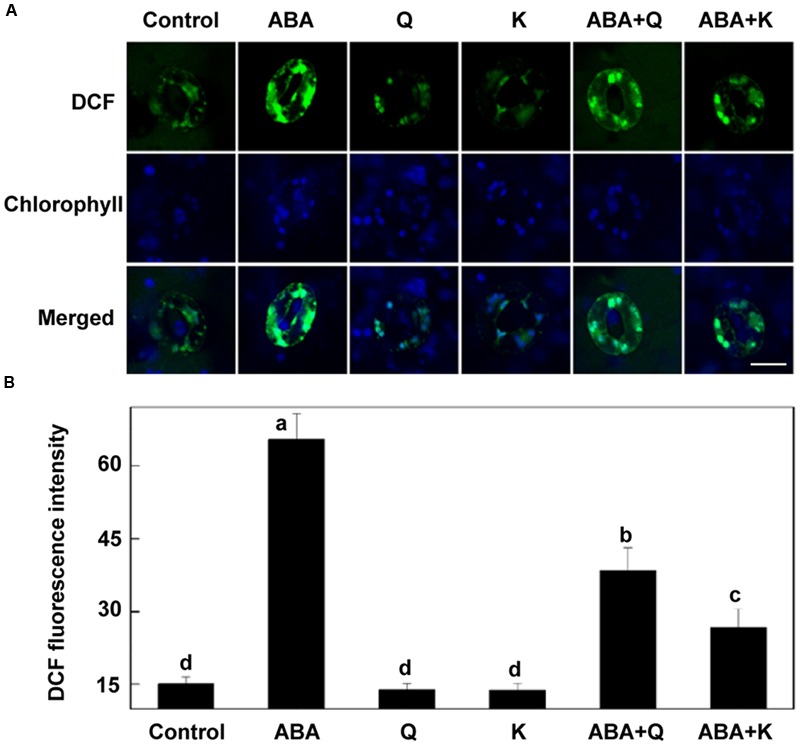
5-aminolevulinic acid (ALA), a new plant growth regulator, can inhibit stomatal closure by reducing H2O2 accumulation in guard cells. Flavonols are a main kind of flavonoids and have been proposed as H2O2 scavengers in guard cells. 5-aminolevulinic acid can significantly improve flavonoids accumulation in plants. However, whether ALA increases flavonols content in guard cells and the role of flavonols in ALA-regulated stomatal movement remains unclear. In this study, we first demonstrated that ALA pretreatment inhibited ABA-induced stomatal closure by reducing H2O2 accumulation in guard cells of Arabidopsis seedlings. This result confirms the inhibitory effect of ALA on stomatal closure and the important role of decreased H2O2 accumulation in this process. We also found that ALA significantly improved flavonols accumulation in guard cells using a flavonol-specific dye. Furthermore, using exogenous quercetin and kaempferol, two major components of flavonols in Arabidopsis leaves, we showed that flavonols accumulation inhibited ABA-induced stomatal movement by suppressing H2O2 in guard cells. Finally, we showed that the inhibitory effect of ALA on ABA-induced stomatal closure was largely impaired in flavonoid-deficient transparent testa4 (tt4) mutant. In addition, exogenous flavonols recovered stomatal responses of tt4 to the wild-type levels. Taken together, we conclude that ALA-induced flavonol accumulation in guard cells is partially involved in the inhibitory effect of ALA on ABA-induced H2O2 accumulation and stomatal closure. Our data provide direct evidence that ALA can regulate stomatal movement by improving flavonols accumulation, revealing new insights into guard cell signaling.
Keywords: 5-aminolevulinic acid (ALA); abscisic acid (ABA); flavonol; hydrogen peroxide; stomatal opening.
Figures




References
-
- Akram N. A., Ashraf M. (2013). Regulation in plant stress tolerance by a potential plant growth regulator, 5-aminolevulinic acid. J. Plant Growth Regul. 32 663–679. 10.1007/s00344-013-9325-9 - DOI
-
- Ali B., Huang C. R., Qi Z. Y., Ali S., Daud M. K., Geng X. X., et al. (2013a). 5-Aminolevulinic acid ameliorates cadmium-induced morphological, biochemical, and ultrastructural changes in seedlings of oilseed rape. Environ. Sci. Pollut. Res. Int. 20 7256–7267. 10.1007/s11356-013-1735-5 - DOI - PubMed
-
- Ali B., Wang B., Ali S., Ghani M. A., Hayat M. T., Yang C., et al. (2013b). 5-Aminolevulinic acid ameliorates the growth, photosynthetic gas exchange capacity, and ultrastructural changes under cadmium stress in Brassica napus L. J. Plant Growth Regul. 32 604–614. 10.1007/s00344-013-9328-6 - DOI
-
- Ali B., Xu X., Gill R. A., Yang S., Ali S., Tahir M., et al. (2014). Promotive role of 5-aminolevulinic acid on mineral nutrients and antioxidative defense system under lead toxicity in Brassica napus. Ind. Crop. Prod. 52 617–626. 10.1016/j.indcrop.2013.11.033 - DOI
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases