

Early generated B1 B cells with restricted BCRs become chronic lymphocytic leukemia with continued c-Myc and low Bmf expression
- PMID: 27899442
- PMCID: PMC5154941
- DOI: 10.1084/jem.20160712
Early generated B1 B cells with restricted BCRs become chronic lymphocytic leukemia with continued c-Myc and low Bmf expression
Abstract
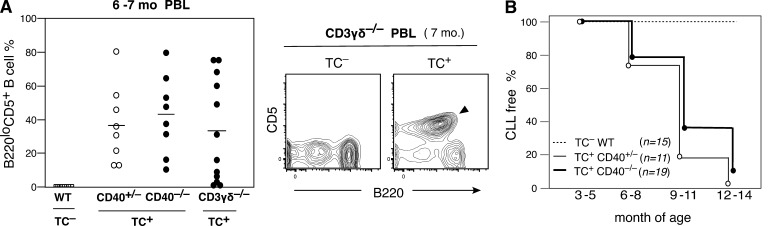
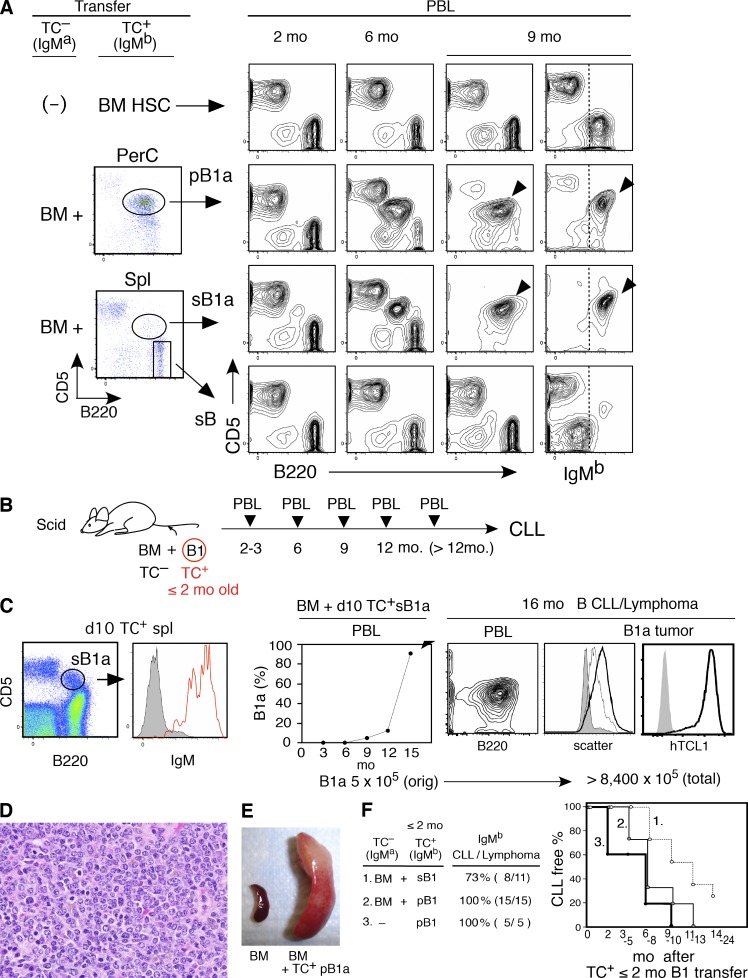
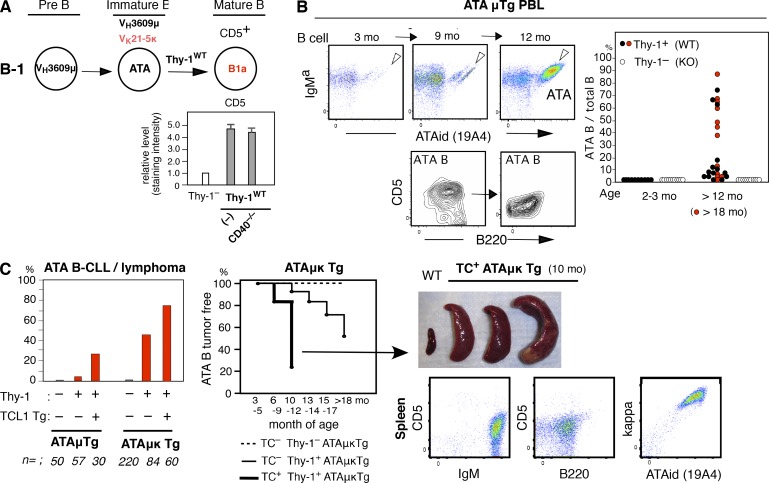
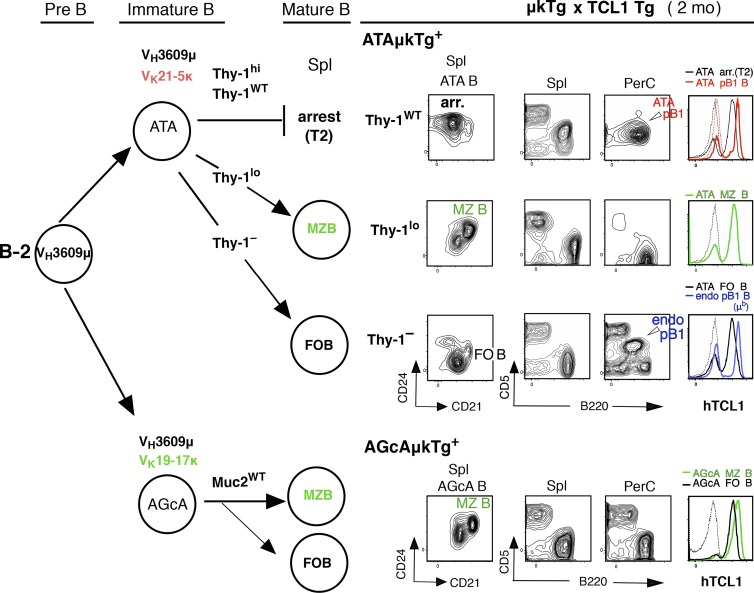
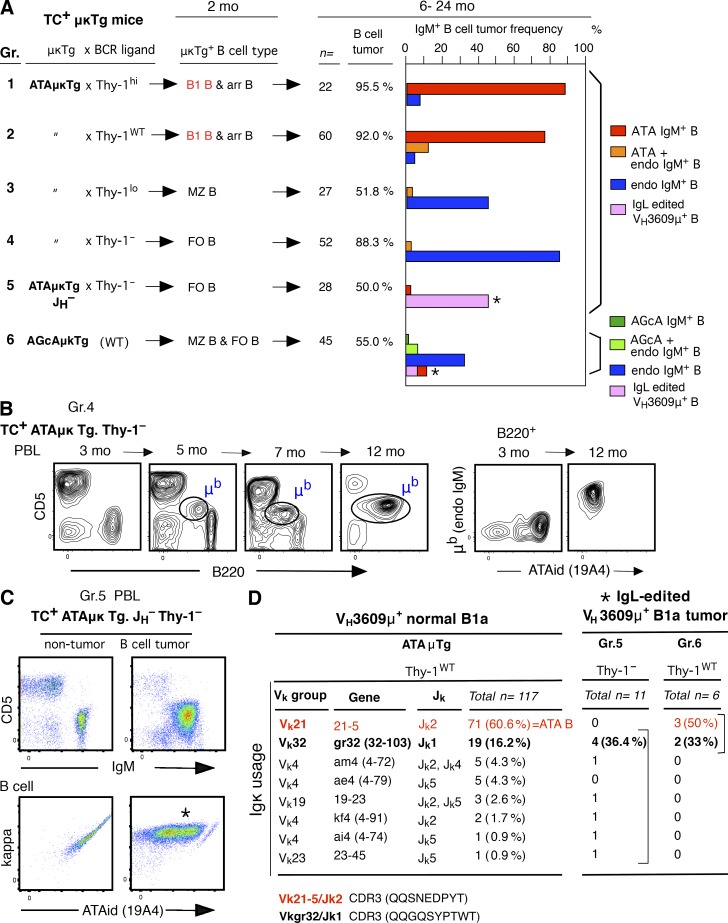
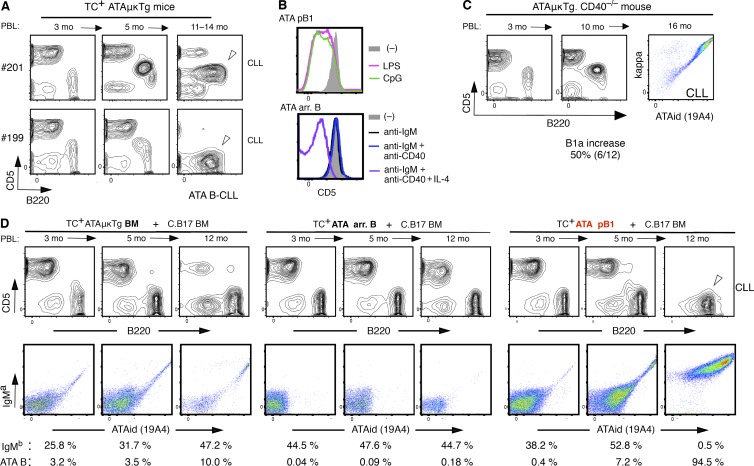
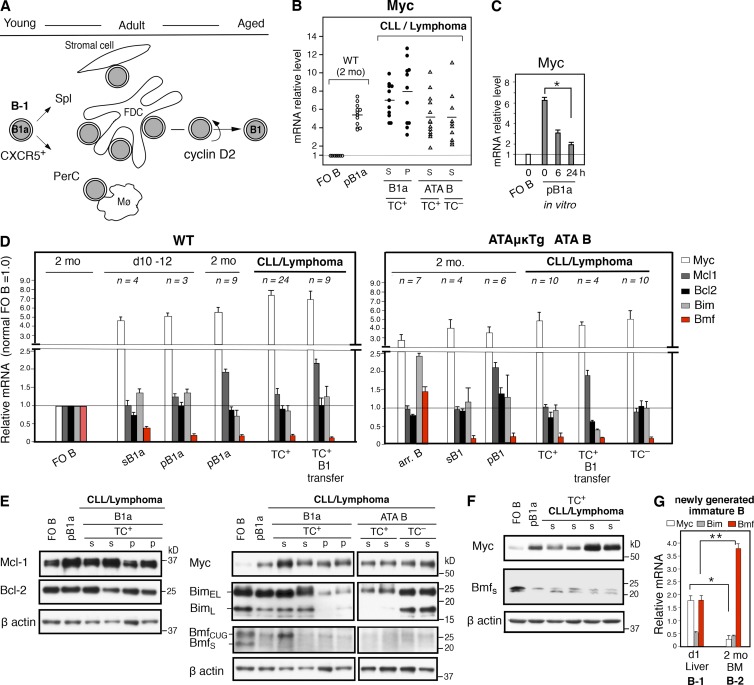
In mice, generation of autoreactive CD5+ B cells occurs as a consequence of BCR signaling induced by (self)-ligand exposure from fetal/neonatal B-1 B cell development. A fraction of these cells self-renew and persist as a minor B1 B cell subset throughout life. Here, we show that transfer of early generated B1 B cells from Eμ-TCL1 transgenic mice resulted in chronic lymphocytic leukemia (CLL) with a biased repertoire, including stereotyped BCRs. Thus, B1 B cells bearing restricted BCRs can become CLL during aging. Increased anti-thymocyte/Thy-1 autoreactive (ATA) BCR cells in the B1 B cell subset by transgenic expression yielded spontaneous ATA B-CLL/lymphoma incidence, enhanced by TCL1 transgenesis. In contrast, ATA B-CLL did not develop from other B cell subsets, even when the identical ATA BCR was expressed on a Thy-1 low/null background. Thus, both a specific BCR and B1 B cell context were important for CLL progression. Neonatal B1 B cells and their CLL progeny in aged mice continued to express moderately up-regulated c-Myc and down-regulated proapoptotic Bmf, unlike most mature B cells in the adult. Thus, there is a genetic predisposition inherent in B-1 development generating restricted BCRs and self-renewal capacity, with both features contributing to potential for progression to CLL.
© 2016 Hayakawa et al.
Figures
References
-
- Bouillet P., Metcalf D., Huang D.C., Tarlinton D.M., Kay T.W., Köntgen F., Adams J.M., and Strasser A.. 1999. Proapoptotic Bcl-2 relative Bim required for certain apoptotic responses, leukocyte homeostasis, and to preclude autoimmunity. Science. 286:1735–1738. 10.1126/science.286.5445.1735 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
