

Whole-transcriptome sequencing identifies a distinct subtype of acute lymphoblastic leukemia with predominant genomic abnormalities of EP300 and CREBBP
- PMID: 27903646
- PMCID: PMC5287225
- DOI: 10.1101/gr.209163.116
Whole-transcriptome sequencing identifies a distinct subtype of acute lymphoblastic leukemia with predominant genomic abnormalities of EP300 and CREBBP
Abstract
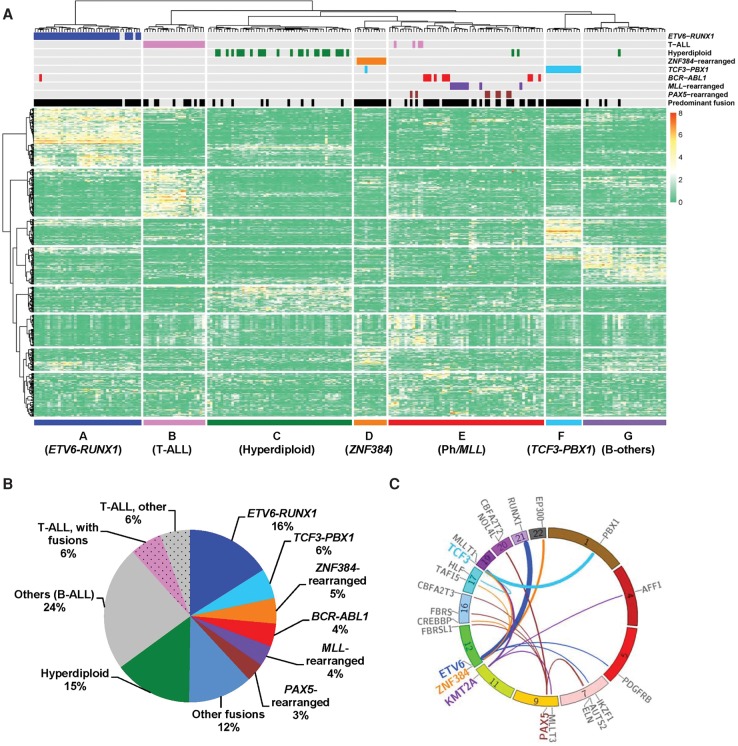
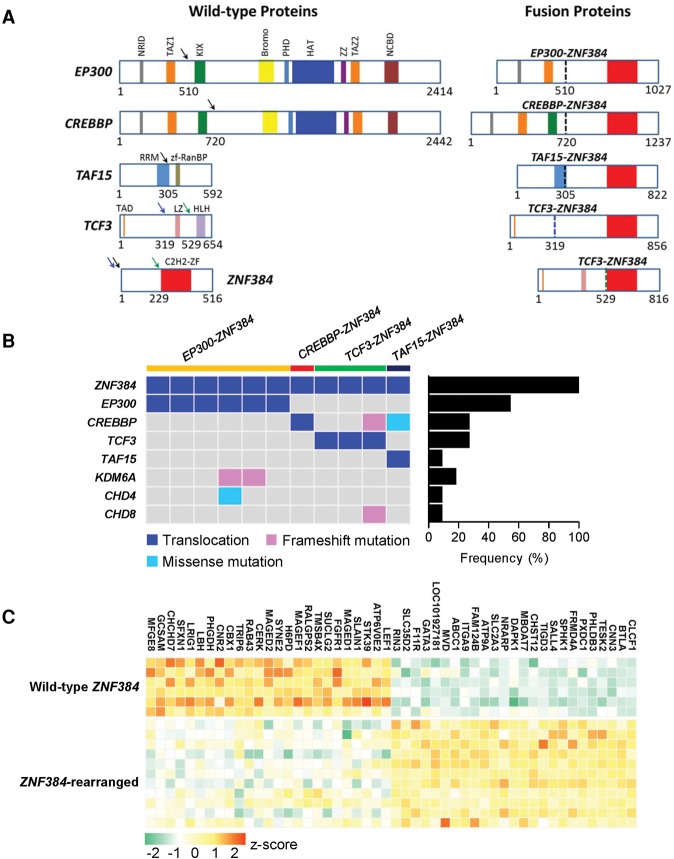
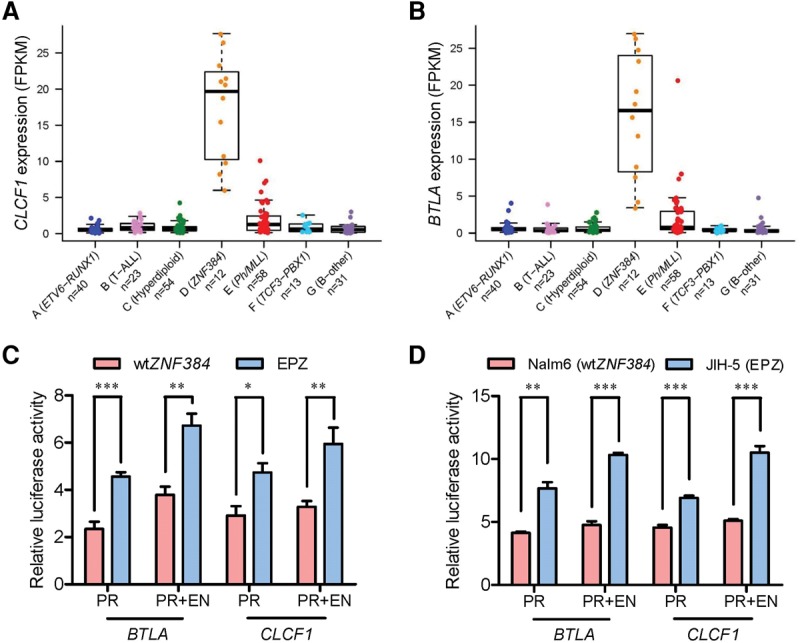
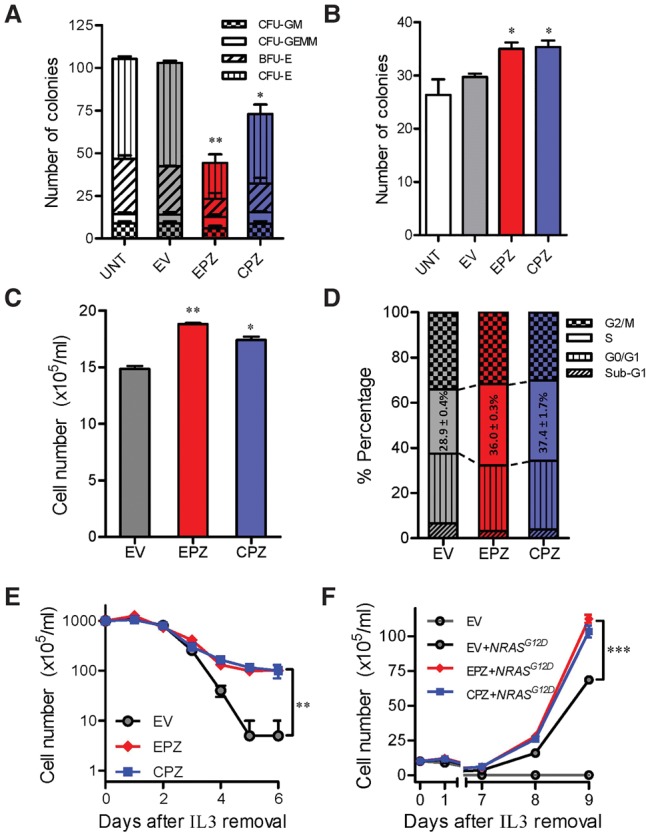
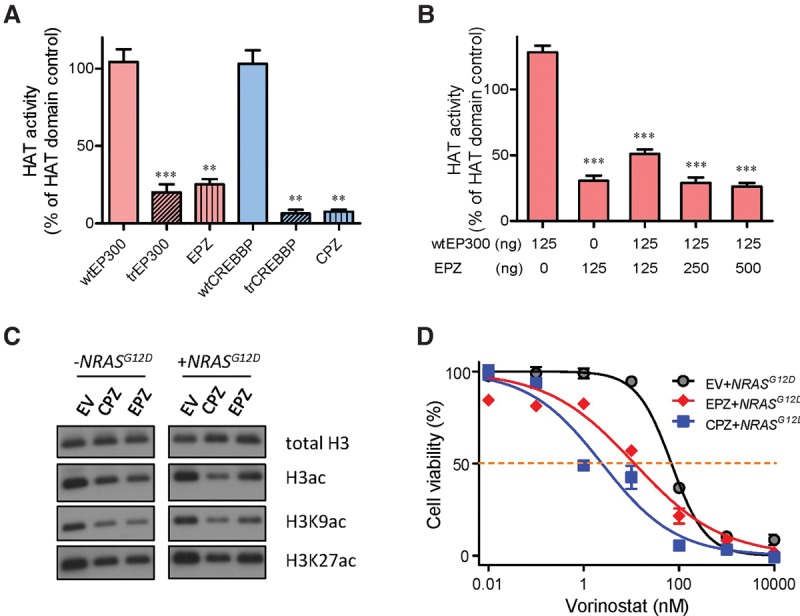
Chromosomal translocations are a genomic hallmark of many hematologic malignancies. Often as initiating events, these structural abnormalities result in fusion proteins involving transcription factors important for hematopoietic differentiation and/or signaling molecules regulating cell proliferation and cell cycle. In contrast, epigenetic regulator genes are more frequently targeted by somatic sequence mutations, possibly as secondary events to further potentiate leukemogenesis. Through comprehensive whole-transcriptome sequencing of 231 children with acute lymphoblastic leukemia (ALL), we identified 58 putative functional and predominant fusion genes in 54.1% of patients (n = 125), 31 of which have not been reported previously. In particular, we described a distinct ALL subtype with a characteristic gene expression signature predominantly driven by chromosomal rearrangements of the ZNF384 gene with histone acetyltransferases EP300 and CREBBP ZNF384-rearranged ALL showed significant up-regulation of CLCF1 and BTLA expression, and ZNF384 fusion proteins consistently showed higher activity to promote transcription of these target genes relative to wild-type ZNF384 in vitro. Ectopic expression of EP300-ZNF384 and CREBBP-ZNF384 fusion altered differentiation of mouse hematopoietic stem and progenitor cells and also potentiated oncogenic transformation in vitro. EP300- and CREBBP-ZNF384 fusions resulted in loss of histone lysine acetyltransferase activity in a dominant-negative fashion, with concomitant global reduction of histone acetylation and increased sensitivity of leukemia cells to histone deacetylase inhibitors. In conclusion, our results indicate that gene fusion is a common class of genomic abnormalities in childhood ALL and that recurrent translocations involving EP300 and CREBBP may cause epigenetic deregulation with potential for therapeutic targeting.
© 2017 Qian et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
References
-
- Ait-Si-Ali S, Ramirez S, Barre FX, Dkhissi F, Magnaghi-Jaulin L, Girault JA, Robin P, Knibiehler M, Pritchard LL, Ducommun B, et al. 1998. Histone acetyltransferase activity of CBP is controlled by cycle-dependent kinases and oncoprotein E1A. Nature 396: 184–186. - PubMed
-
- Anderson K, Lutz C, van Delft FW, Bateman CM, Guo Y, Colman SM, Kempski H, Moorman AV, Titley I, Swansbury J, et al. 2011. Genetic variegation of clonal architecture and propagating cells in leukaemia. Nature 469: 356–361. - PubMed
-
- Atak ZK, Gianfelici V, Hulselmans G, De Keersmaecker K, Devasia AG, Geerdens E, Mentens N, Chiaretti S, Durinck K, Uyttebroeck A, et al. 2013. Comprehensive analysis of transcriptome variation uncovers known and novel driver events in T-cell acute lymphoblastic leukemia. PLoS Genet 9: e1003997. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous