

The PLK4-STIL-SAS-6 module at the core of centriole duplication
- PMID: 27911707
- PMCID: PMC5095913
- DOI: 10.1042/BST20160116
The PLK4-STIL-SAS-6 module at the core of centriole duplication
Abstract
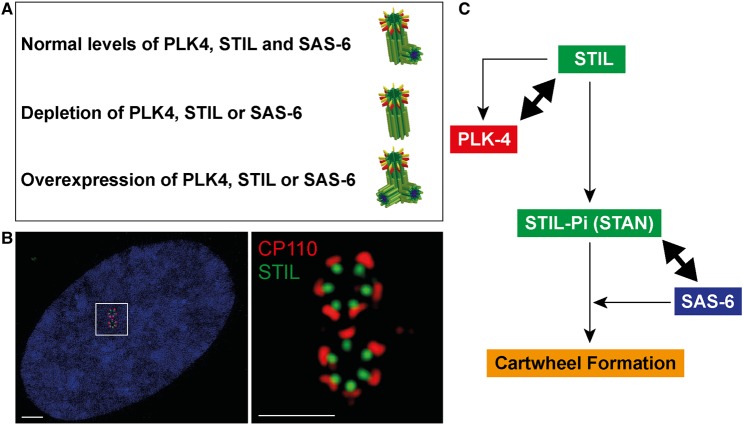
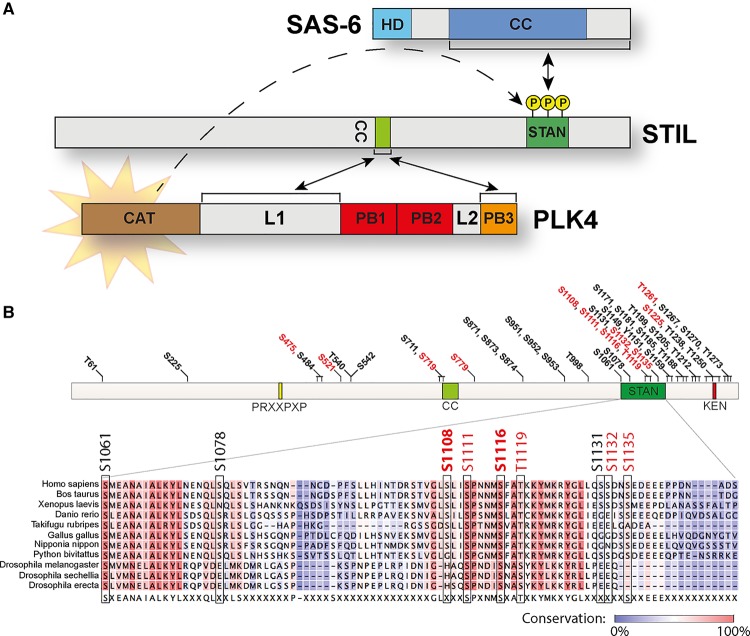
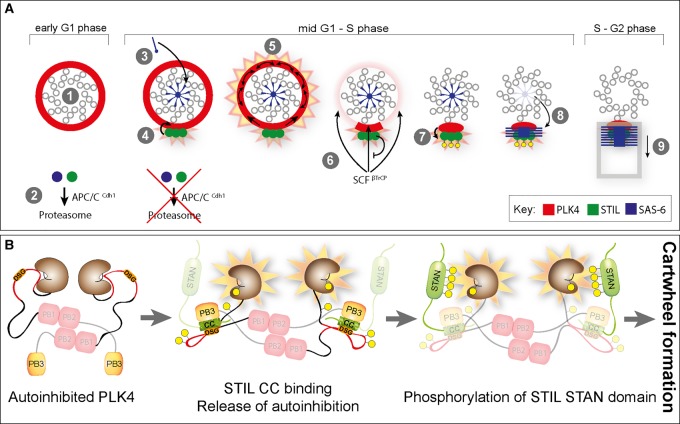
Centrioles are microtubule-based core components of centrosomes and cilia. They are duplicated exactly once during S-phase progression. Central to formation of each new (daughter) centriole is the formation of a nine-fold symmetrical cartwheel structure onto which microtubule triplets are deposited. In recent years, a module comprising the protein kinase polo-like kinase 4 (PLK4) and the two proteins STIL and SAS-6 have been shown to stay at the core of centriole duplication. Depletion of any one of these three proteins blocks centriole duplication and, conversely, overexpression causes centriole amplification. In this short review article, we summarize recent insights into how PLK4, STIL and SAS-6 co-operate in space and time to form a new centriole. These advances begin to shed light on the very first steps of centriole biogenesis.
Keywords: PLK4; SAS-6; cartwheel formation; centriole duplication; centrosomes.
© 2016 The Author(s).
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources