

Two crystal structures reveal design for repurposing the C-Ala domain of human AlaRS
- PMID: 27911835
- PMCID: PMC5167178
- DOI: 10.1073/pnas.1617316113
Two crystal structures reveal design for repurposing the C-Ala domain of human AlaRS
Abstract
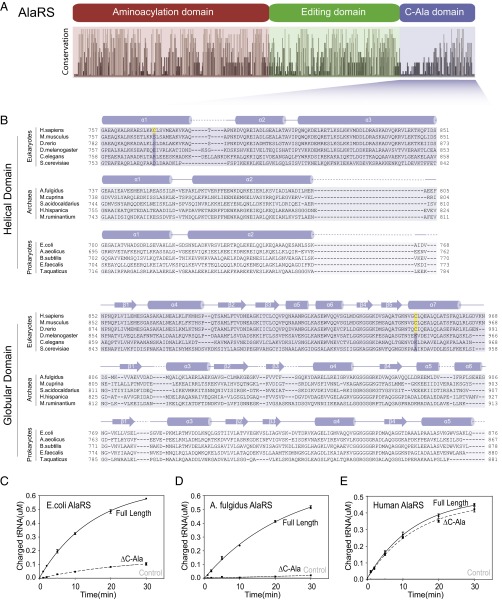
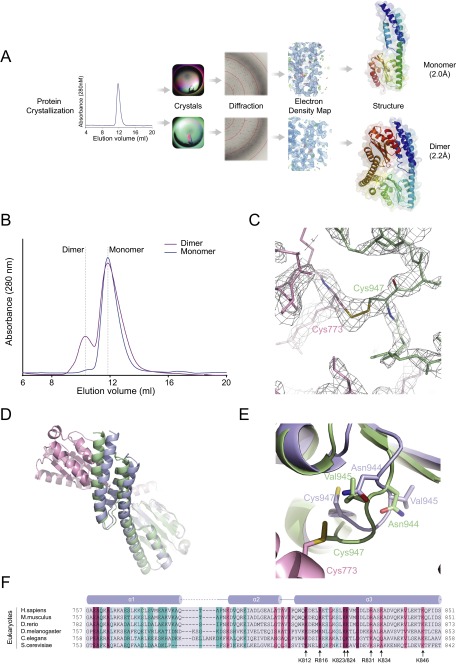
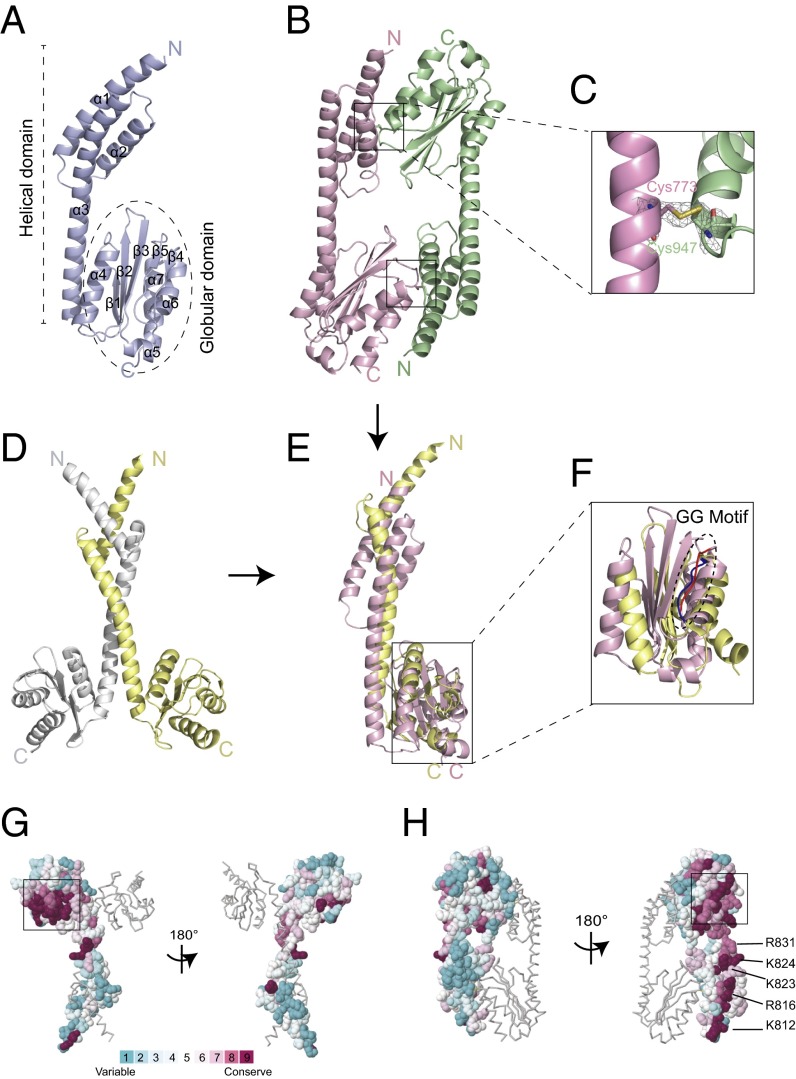
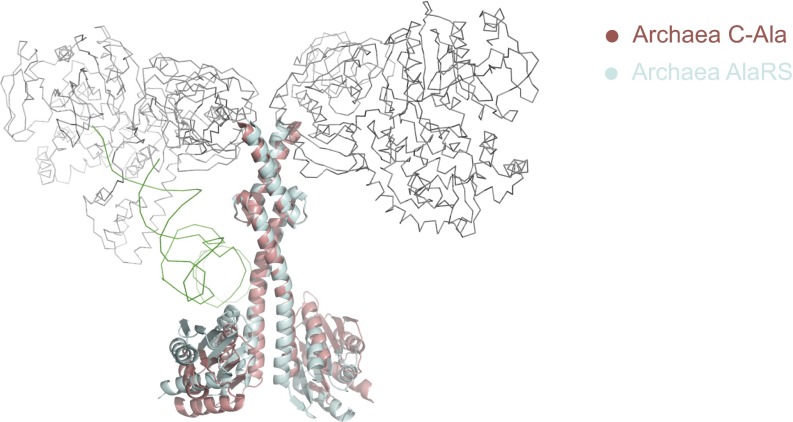
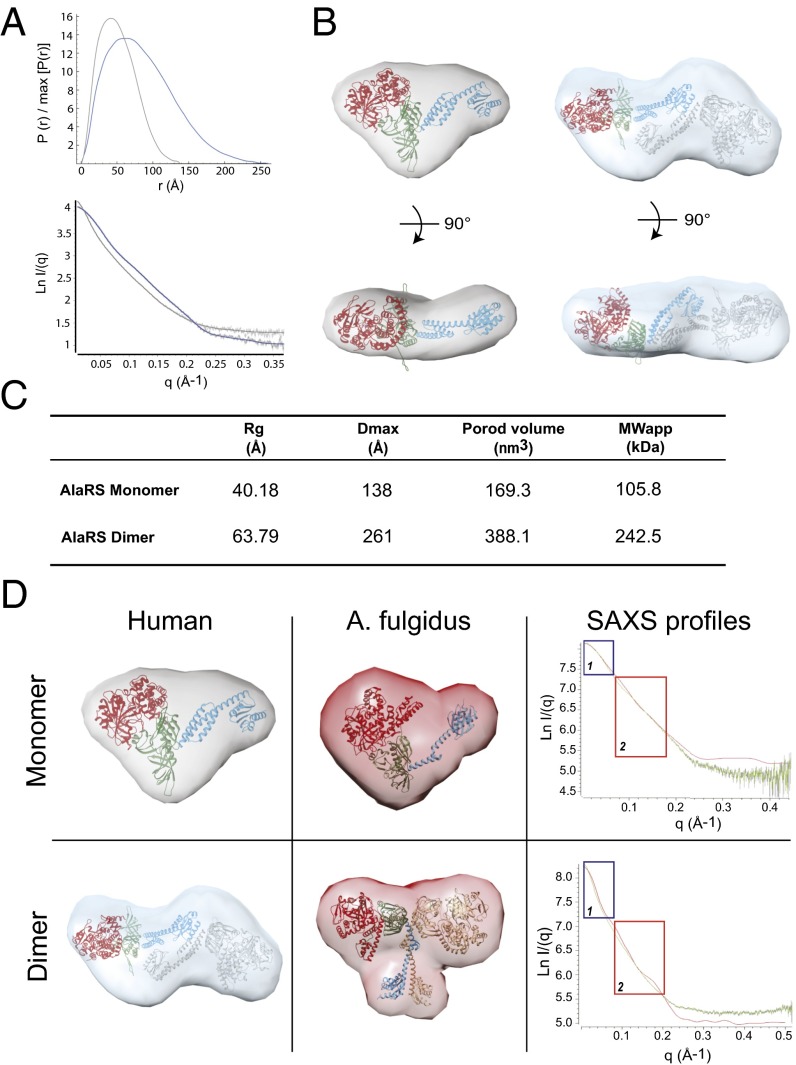
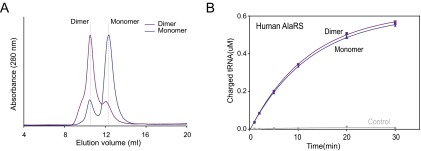
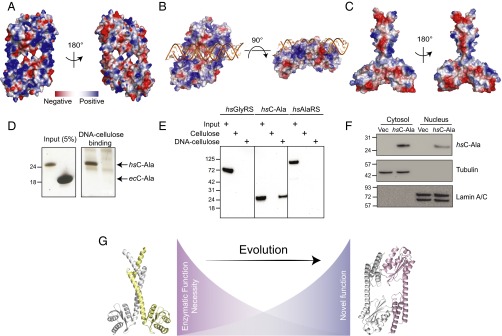
The 20 aminoacyl tRNA synthetases (aaRSs) couple each amino acid to their cognate tRNAs. During evolution, 19 aaRSs expanded by acquiring novel noncatalytic appended domains, which are absent from bacteria and many lower eukaryotes but confer extracellular and nuclear functions in higher organisms. AlaRS is the single exception, with an appended C-terminal domain (C-Ala) that is conserved from prokaryotes to humans but with a wide sequence divergence. In human cells, C-Ala is also a splice variant of AlaRS. Crystal structures of two forms of human C-Ala, and small-angle X-ray scattering of AlaRS, showed that the large sequence divergence of human C-Ala reshaped C-Ala in a way that changed the global architecture of AlaRS. This reshaping removes the role of C-Ala in prokaryotes for docking tRNA and instead repurposes it to form a dimer interface presenting a DNA-binding groove. This groove cannot form with the bacterial ortholog. Direct DNA binding by human C-Ala, but not by bacterial C-Ala, was demonstrated. Thus, instead of acquiring a novel appended domain like other human aaRSs, which engendered novel functions, a new AlaRS architecture was created by diversifying a preexisting appended domain.
Keywords: DNA binding; appended domain; evolution; splice variant; structural plasticity.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
