

Conflict Resolution in the Genome: How Transcription and Replication Make It Work
- PMID: 27912056
- PMCID: PMC5141617
- DOI: 10.1016/j.cell.2016.09.053
Conflict Resolution in the Genome: How Transcription and Replication Make It Work
Abstract
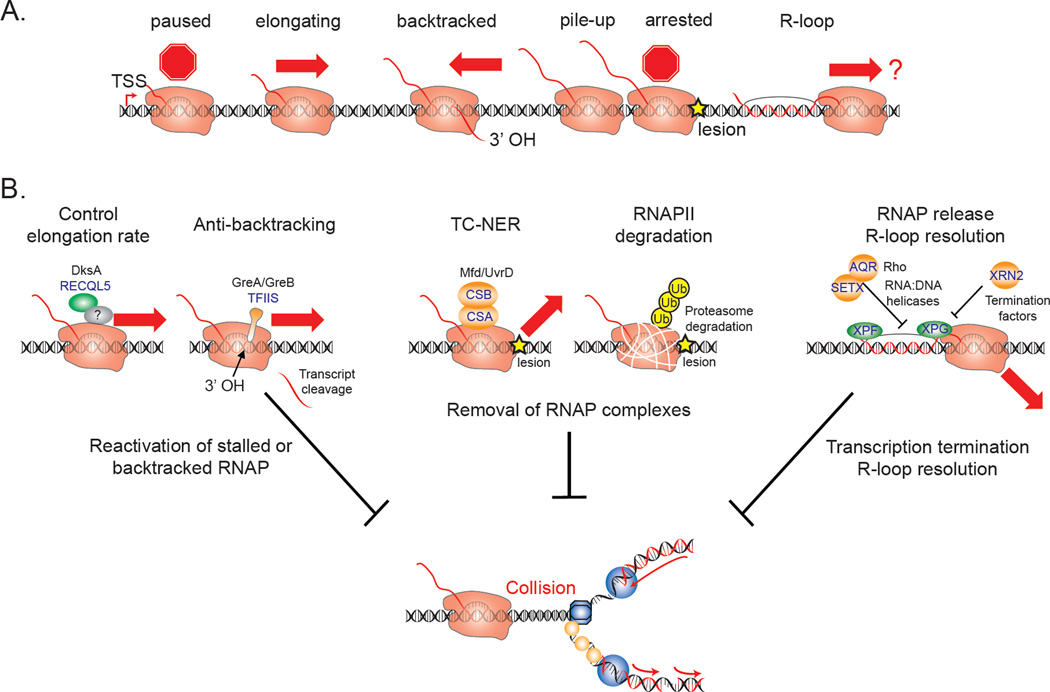
The complex machineries involved in replication and transcription translocate along the same DNA template, often in opposing directions and at different rates. These processes routinely interfere with each other in prokaryotes, and mounting evidence now suggests that RNA polymerase complexes also encounter replication forks in higher eukaryotes. Indeed, cells rely on numerous mechanisms to avoid, tolerate, and resolve such transcription-replication conflicts, and the absence of these mechanisms can lead to catastrophic effects on genome stability and cell viability. In this article, we review the cellular responses to transcription-replication conflicts and highlight how these inevitable encounters shape the genome and impact diverse cellular processes.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures



References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
