

Replication stalling and DNA microsatellite instability
- PMID: 27914716
- PMCID: PMC5440219
- DOI: 10.1016/j.bpc.2016.11.007
Replication stalling and DNA microsatellite instability
Abstract
Microsatellites are short, tandemly repeated DNA motifs of 1-6 nucleotides, also termed simple sequence repeats (SRSs) or short tandem repeats (STRs). Collectively, these repeats comprise approximately 3% of the human genome Subramanian et al. (2003), Lander and Lander (2001) [1,2], and represent a large reservoir of loci highly prone to mutations Sun et al. (2012), Ellegren (2004) [3,4] that contribute to human evolution and disease. Microsatellites are known to stall and reverse replication forks in model systems Pelletier et al. (2003), Samadashwily et al. (1997), Kerrest et al. (2009) [5-7], and are hotspots of chromosomal double strand breaks (DSBs). We briefly review the relationship of these repeated sequences to replication stalling and genome instability, and present recent data on the impact of replication stress on DNA fragility at microsatellites in vivo.
Copyright © 2016 Elsevier B.V. All rights reserved.
Figures

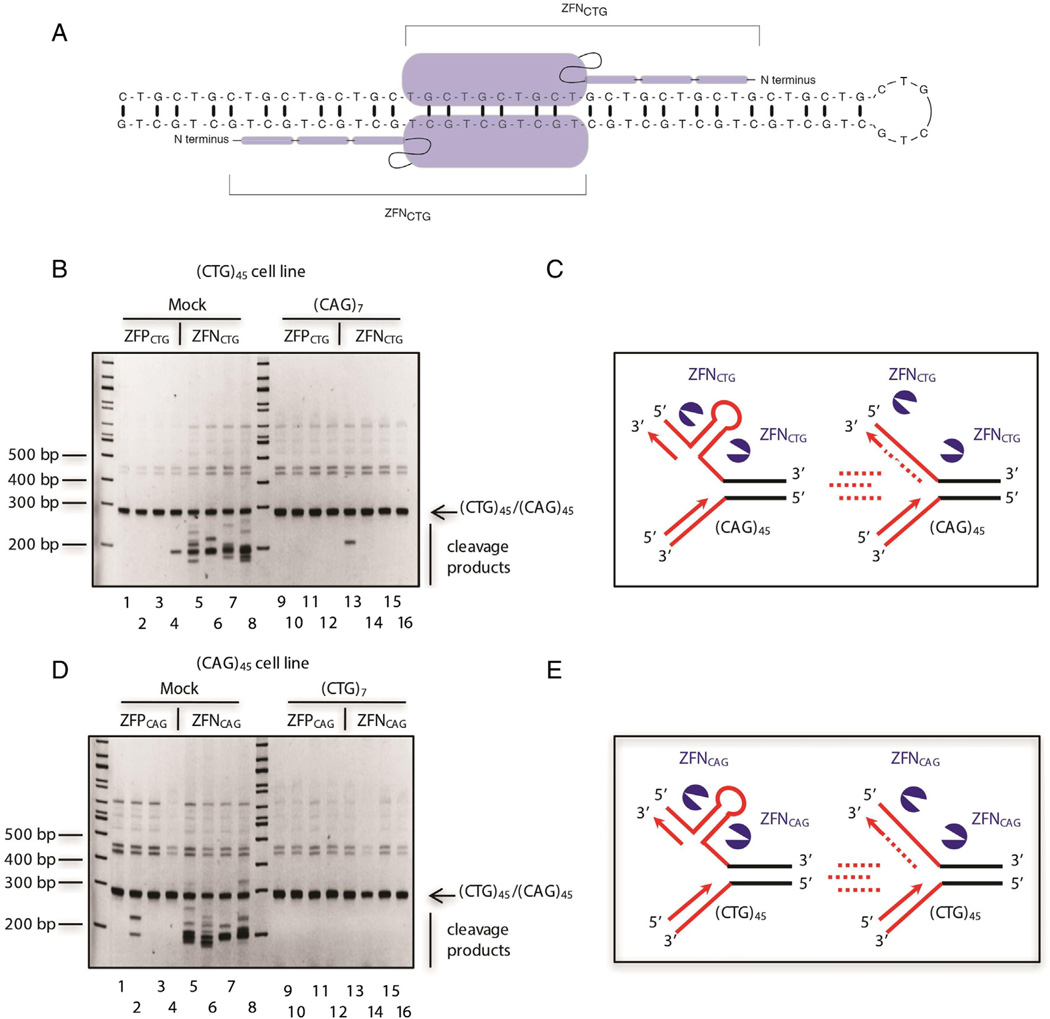


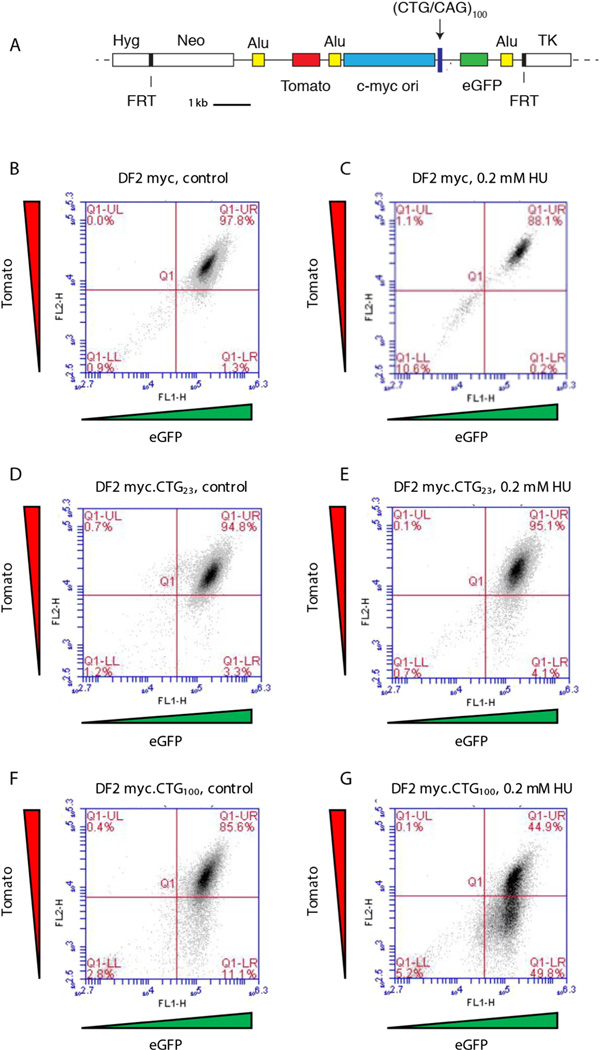
Similar articles
-
Replication stress at microsatellites causes DNA double-strand breaks and break-induced replication.J Biol Chem. 2020 Nov 6;295(45):15378-15397. doi: 10.1074/jbc.RA120.013495. Epub 2020 Sep 1. J Biol Chem. 2020. PMID: 32873711 Free PMC article.
-
Replication of [AT/TA]25 Microsatellite Sequences by Human DNA Polymerase δ Holoenzymes Is Dependent on dNTP and RPA Levels.Biochemistry. 2024 Apr 16;63(8):969-983. doi: 10.1021/acs.biochem.4c00006. Epub 2024 Mar 26. Biochemistry. 2024. PMID: 38623046 Free PMC article.
-
Genome-wide Identification of Structure-Forming Repeats as Principal Sites of Fork Collapse upon ATR Inhibition.Mol Cell. 2018 Oct 18;72(2):222-238.e11. doi: 10.1016/j.molcel.2018.08.047. Epub 2018 Oct 4. Mol Cell. 2018. PMID: 30293786 Free PMC article.
-
Break-induced replication links microsatellite expansion to complex genome rearrangements.Bioessays. 2017 Aug;39(8):10.1002/bies.201700025. doi: 10.1002/bies.201700025. Epub 2017 Jun 16. Bioessays. 2017. PMID: 28621832 Free PMC article. Review.
-
Evolutionary dynamics of microsatellite DNA.Chromosoma. 2000 Sep;109(6):365-71. doi: 10.1007/s004120000089. Chromosoma. 2000. PMID: 11072791 Review.
Cited by
-
Restarted replication forks are error-prone and cause CAG repeat expansions and contractions.PLoS Genet. 2021 Oct 21;17(10):e1009863. doi: 10.1371/journal.pgen.1009863. eCollection 2021 Oct. PLoS Genet. 2021. PMID: 34673780 Free PMC article.
-
Genetic diversity and population structure of Aedes aegypti after massive vector control for dengue fever prevention in Yunnan border areas.Sci Rep. 2020 Jul 29;10(1):12731. doi: 10.1038/s41598-020-69668-7. Sci Rep. 2020. PMID: 32728176 Free PMC article.
-
A Long Polymorphic GT Microsatellite within a Gene Promoter Mediates Non-Imprinted Allele-Specific DNA Methylation of a CpG Island in a Goldfish Inter-Strain Hybrid.Int J Mol Sci. 2019 Aug 12;20(16):3923. doi: 10.3390/ijms20163923. Int J Mol Sci. 2019. PMID: 31409051 Free PMC article.
-
A comparative cytogenetic study of Hypsibarbusmalcolmi and H.wetmorei (Cyprinidae, Poropuntiini).Comp Cytogenet. 2023 Sep 15;17:181-194. doi: 10.3897/compcytogen.17.107703. eCollection 2023. Comp Cytogenet. 2023. PMID: 37794860 Free PMC article.
-
Homologous recombination occurs frequently at innate GT microsatellites in normal somatic and germ cells in vivo.BMC Genomics. 2018 May 11;19(1):359. doi: 10.1186/s12864-018-4758-y. BMC Genomics. 2018. PMID: 29751739 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
