

The Gene Regulatory Network of Lens Induction Is Wired through Meis-Dependent Shadow Enhancers of Pax6
- PMID: 27918583
- PMCID: PMC5137874
- DOI: 10.1371/journal.pgen.1006441
The Gene Regulatory Network of Lens Induction Is Wired through Meis-Dependent Shadow Enhancers of Pax6
Abstract
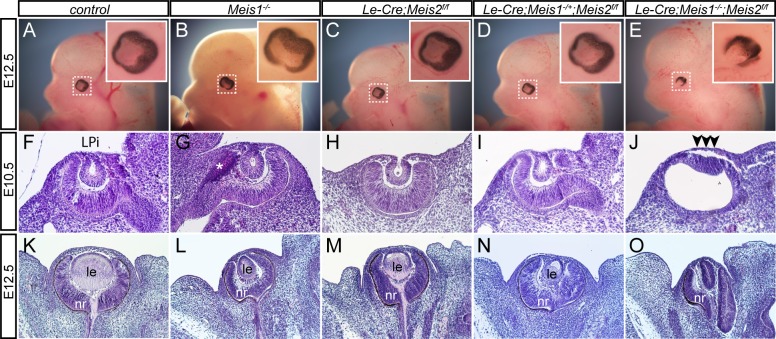
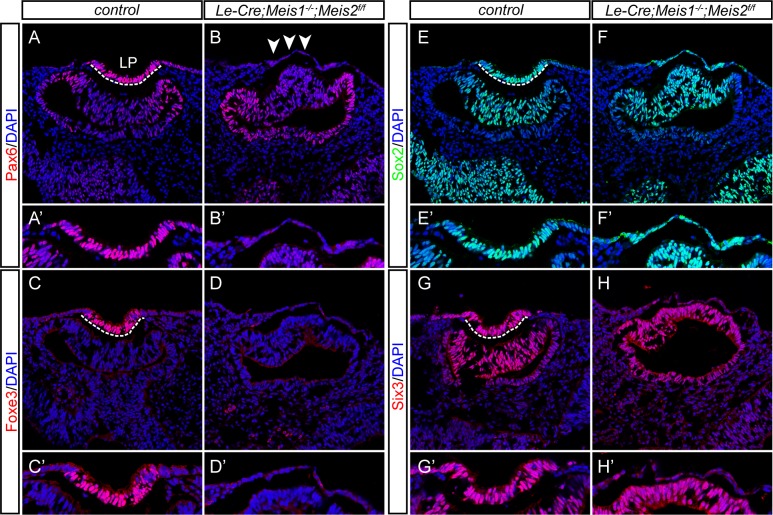
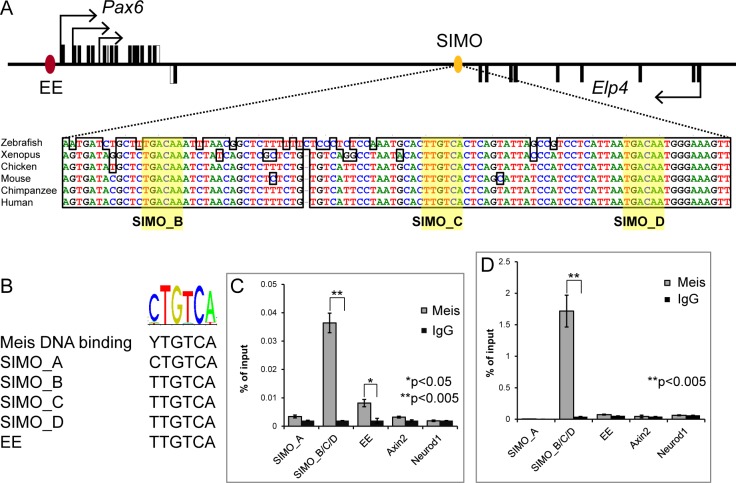
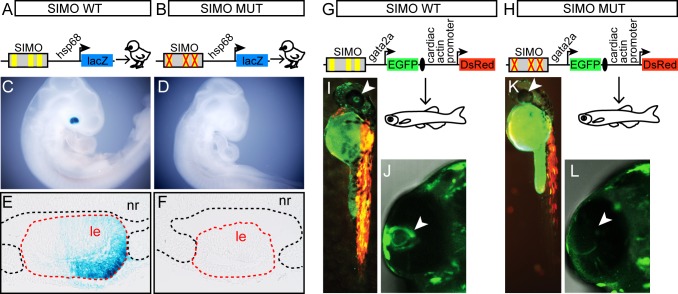
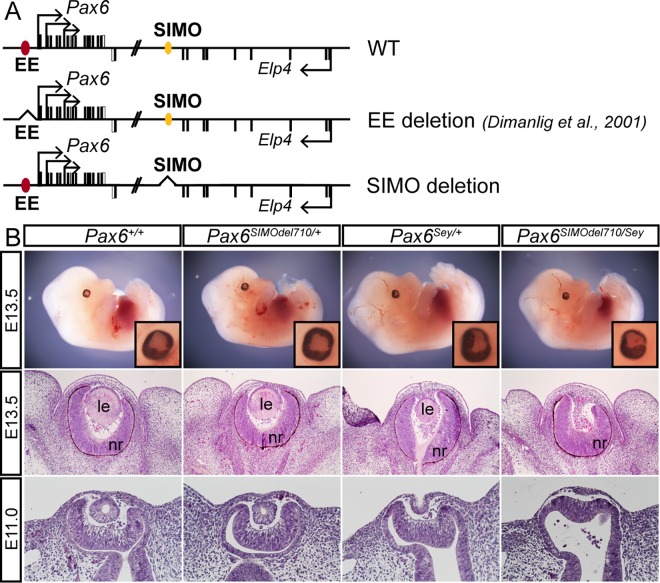
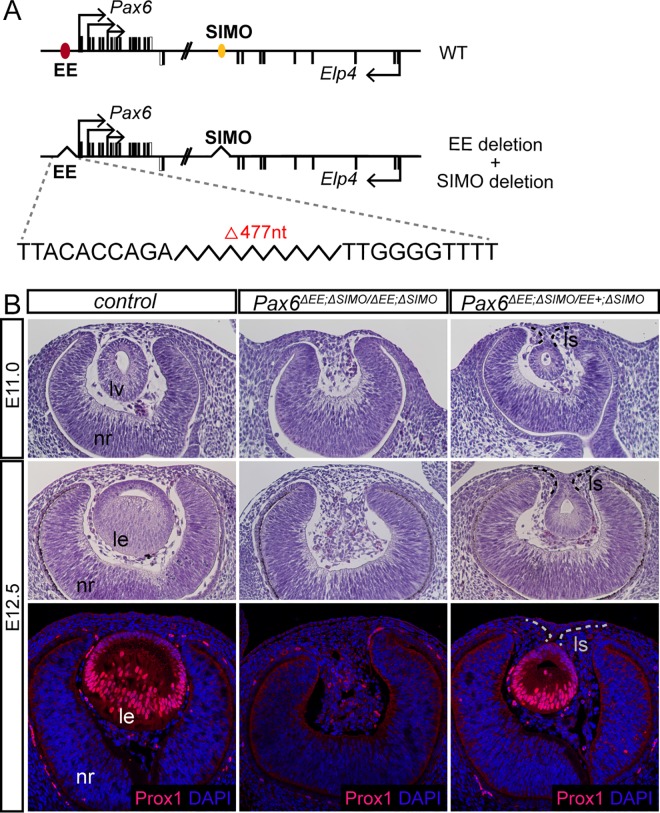
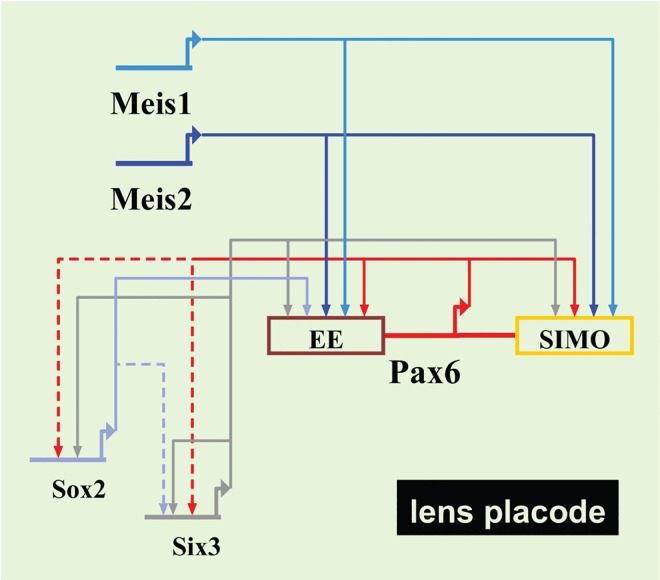
Lens induction is a classical developmental model allowing investigation of cell specification, spatiotemporal control of gene expression, as well as how transcription factors are integrated into highly complex gene regulatory networks (GRNs). Pax6 represents a key node in the gene regulatory network governing mammalian lens induction. Meis1 and Meis2 homeoproteins are considered as essential upstream regulators of Pax6 during lens morphogenesis based on their interaction with the ectoderm enhancer (EE) located upstream of Pax6 transcription start site. Despite this generally accepted regulatory pathway, Meis1-, Meis2- and EE-deficient mice have surprisingly mild eye phenotypes at placodal stage of lens development. Here, we show that simultaneous deletion of Meis1 and Meis2 in presumptive lens ectoderm results in arrested lens development in the pre-placodal stage, and neither lens placode nor lens is formed. We found that in the presumptive lens ectoderm of Meis1/Meis2 deficient embryos Pax6 expression is absent. We demonstrate using chromatin immunoprecipitation (ChIP) that in addition to EE, Meis homeoproteins bind to a remote, ultraconserved SIMO enhancer of Pax6. We further show, using in vivo gene reporter analyses, that the lens-specific activity of SIMO enhancer is dependent on the presence of three Meis binding sites, phylogenetically conserved from man to zebrafish. Genetic ablation of EE and SIMO enhancers demostrates their requirement for lens induction and uncovers an apparent redundancy at early stages of lens development. These findings identify a genetic requirement for Meis1 and Meis2 during the early steps of mammalian eye development. Moreover, they reveal an apparent robustness in the gene regulatory mechanism whereby two independent "shadow enhancers" maintain critical levels of a dosage-sensitive gene, Pax6, during lens induction.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Gunhaga L. The lens: a classical model of embryonic induction providing new insights into cell determination in early development. Philosophical transactions of the Royal Society of London Series B, Biological sciences. 2011;366(1568):1193–203. PubMed Central PMCID: PMC3061103. 10.1098/rstb.2010.0175 - DOI - PMC - PubMed
-
- Hogan BL, Horsburgh G, Cohen J, Hetherington CM, Fisher G, Lyon MF. Small eyes (Sey): a homozygous lethal mutation on chromosome 2 which affects the differentiation of both lens and nasal placodes in the mouse. Journal of embryology and experimental morphology. 1986;97:95–110. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
