

SUPPRESSOR OF FRIGIDA (SUF4) Supports Gamete Fusion via Regulating Arabidopsis EC1 Gene Expression
- PMID: 27920160
- PMCID: PMC5210714
- DOI: 10.1104/pp.16.01024
SUPPRESSOR OF FRIGIDA (SUF4) Supports Gamete Fusion via Regulating Arabidopsis EC1 Gene Expression
Abstract
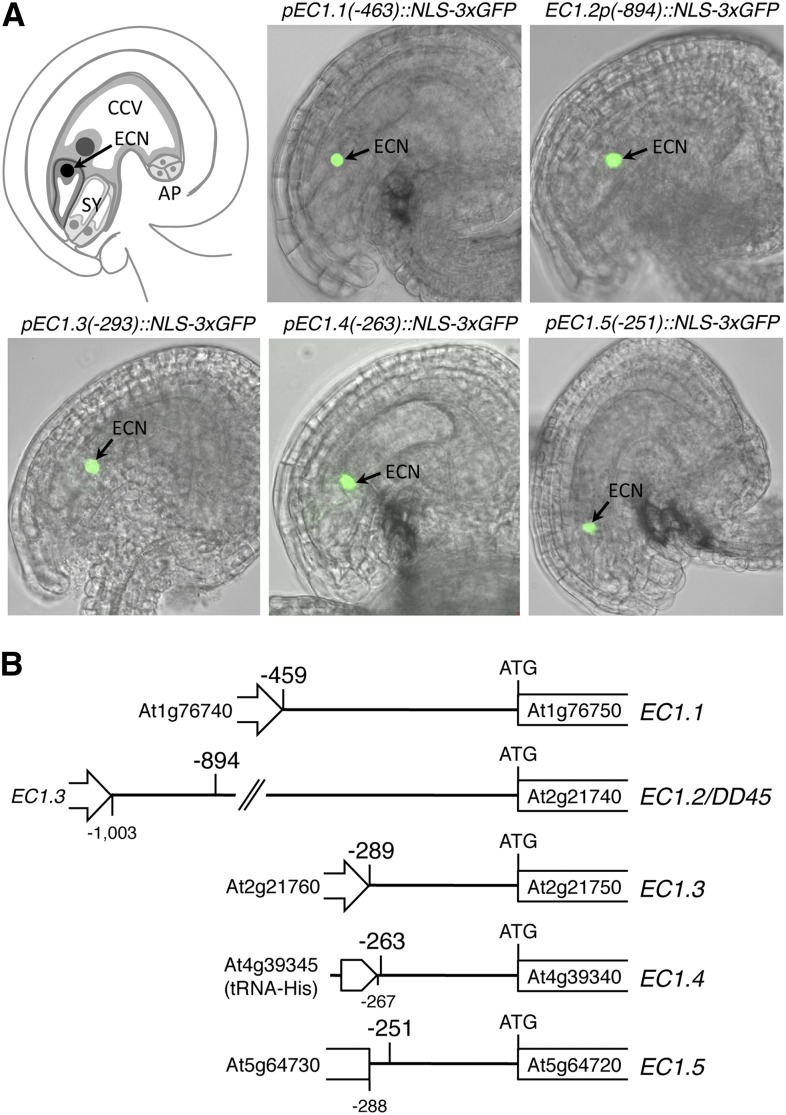
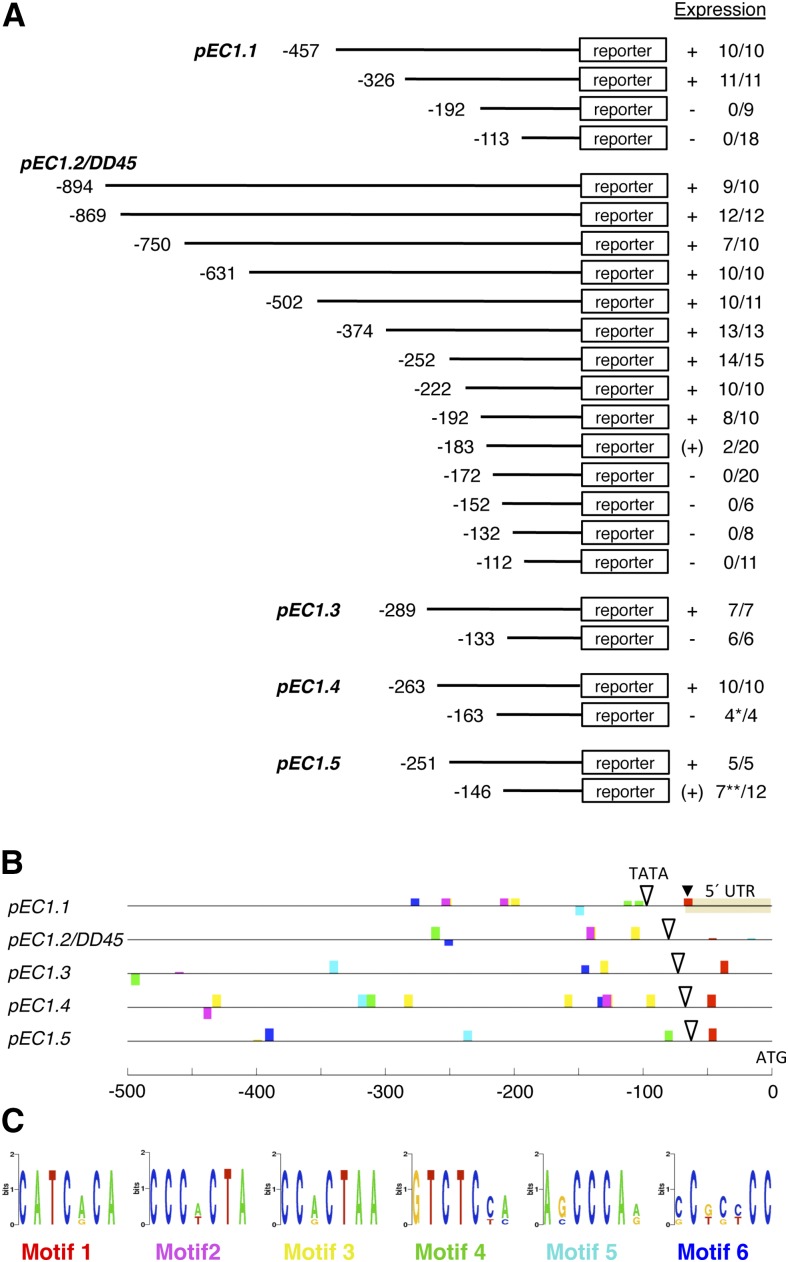
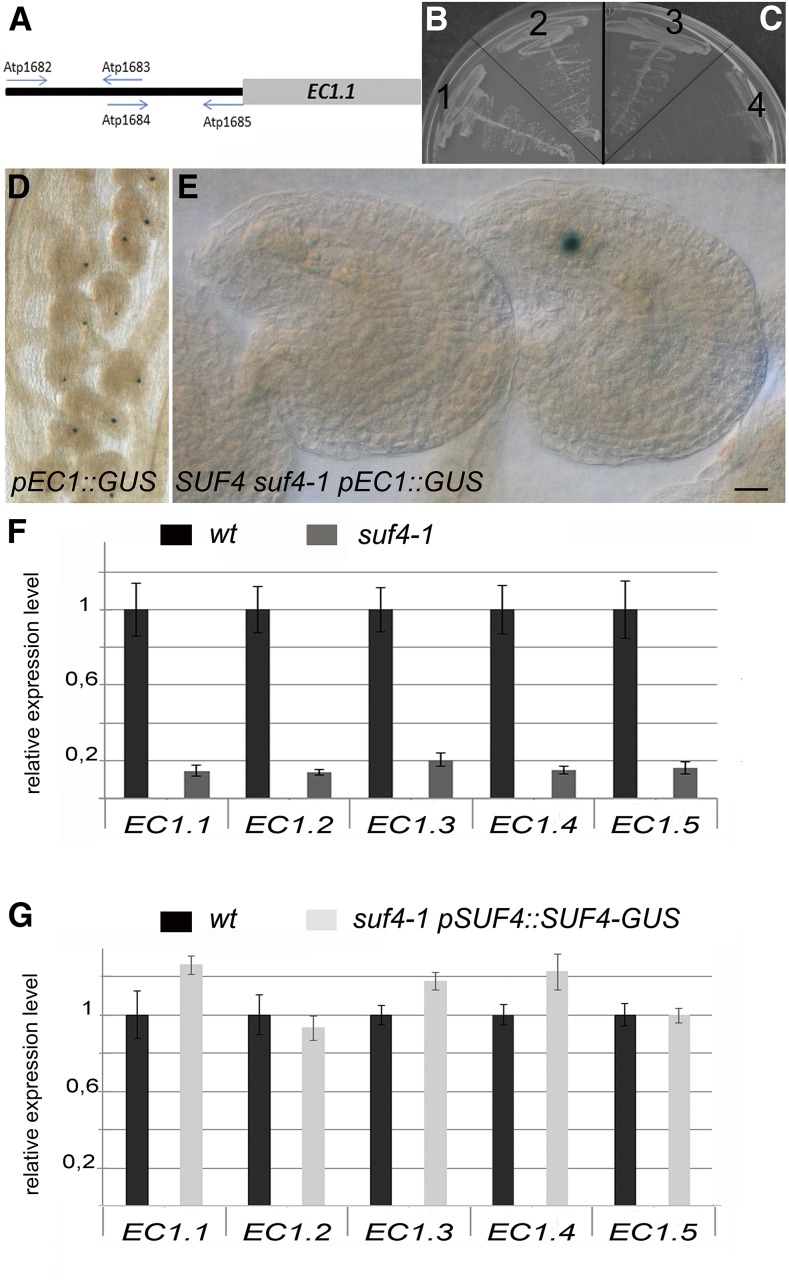
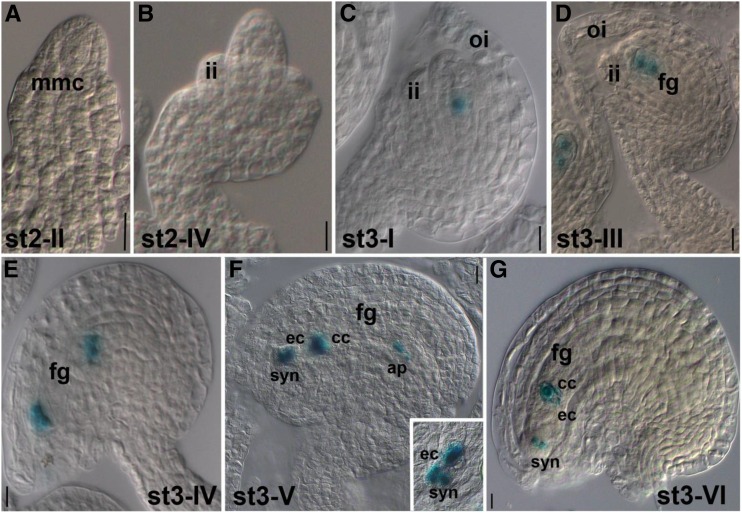
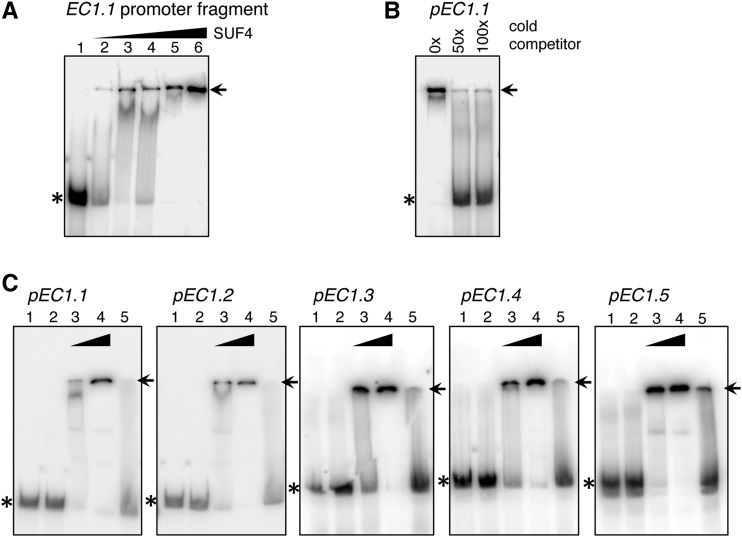
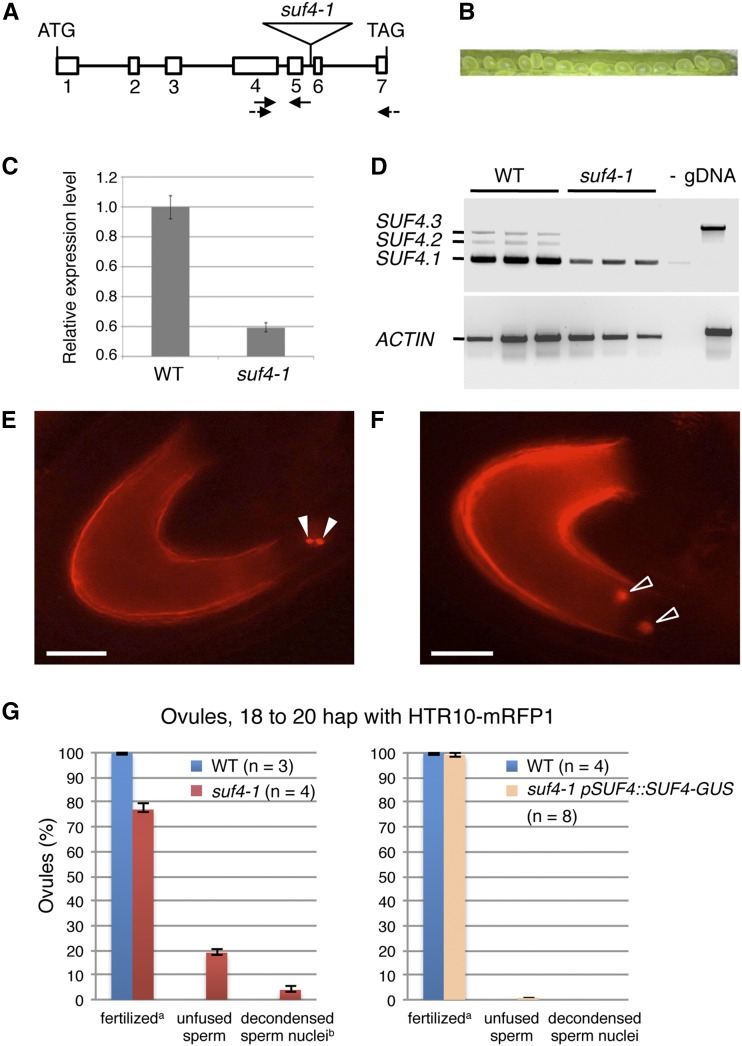
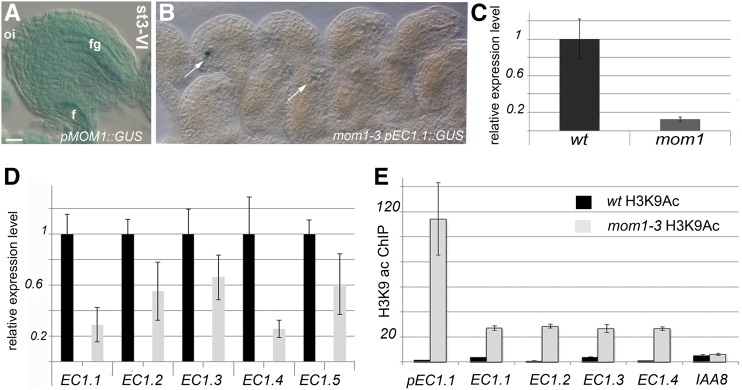
The EGG CELL1 (EC1) gene family of Arabidopsis (Arabidopsis thaliana) comprises five members that are specifically expressed in the egg cell and redundantly control gamete fusion during double fertilization. We investigated the activity of all five EC1 promoters in promoter-deletion studies and identified SUF4 (SUPPRESSOR OF FRIGIDA4), a C2H2 transcription factor, as a direct regulator of the EC1 gene expression. In particular, we demonstrated that SUF4 binds to all five Arabidopsis EC1 promoters, thus regulating their expression. The down-regulation of SUF4 in homozygous suf4-1 ovules results in reduced EC1 expression and delayed sperm fusion, which can be rescued by expressing SUF4-β-glucuronidase under the control of the SUF4 promoter. To identify more gene products able to regulate EC1 expression together with SUF4, we performed coexpression studies that led to the identification of MOM1 (MORPHEUS' MOLECULE1), a component of a silencing mechanism that is independent of DNA methylation marks. In mom1-3 ovules, both SUF4 and EC1 genes are down-regulated, and EC1 genes show higher levels of histone 3 lysine-9 acetylation, suggesting that MOM1 contributes to the regulation of SUF4 and EC1 gene expression.
© 2017 American Society of Plant Biologists. All Rights Reserved.
Figures
Similar articles
-
SUPPRESSOR OF FRIGIDA4, encoding a C2H2-Type zinc finger protein, represses flowering by transcriptional activation of Arabidopsis FLOWERING LOCUS C.Plant Cell. 2006 Nov;18(11):2985-98. doi: 10.1105/tpc.106.045179. Epub 2006 Nov 30. Plant Cell. 2006. PMID: 17138694 Free PMC article.
-
SUPPRESSOR OF FRIGIDA 4 cooperates with the histone methylation reader EBS to positively regulate root development.Plant Physiol. 2024 Dec 2;196(4):2199-2212. doi: 10.1093/plphys/kiae321. Plant Physiol. 2024. PMID: 38875008
-
Egg cell-secreted EC1 triggers sperm cell activation during double fertilization.Science. 2012 Nov 23;338(6110):1093-7. doi: 10.1126/science.1223944. Science. 2012. PMID: 23180860
-
Same same but different: sperm-activating EC1 and ECA1 gametogenesis-related family proteins.Biochem Soc Trans. 2014 Apr;42(2):401-7. doi: 10.1042/BST20140039. Biochem Soc Trans. 2014. PMID: 24646251 Review.
-
Epigenetic silencing of endogenous repetitive sequences by MORPHEUS' MOLECULE1 in Arabidopsis thaliana.Epigenetics. 2010 Oct 1;5(7):562-5. doi: 10.4161/epi.5.7.12518. Epigenetics. 2010. PMID: 21464619 Review.
Cited by
-
Time-Course Transcriptome Analysis of Arabidopsis Siliques Discloses Genes Essential for Fruit Development and Maturation.Plant Physiol. 2018 Nov;178(3):1249-1268. doi: 10.1104/pp.18.00727. Epub 2018 Oct 1. Plant Physiol. 2018. PMID: 30275057 Free PMC article.
-
Trans-splicing of plastid rps12 transcripts, mediated by AtPPR4, is essential for embryo patterning in Arabidopsis thaliana.Planta. 2018 Jul;248(1):257-265. doi: 10.1007/s00425-018-2896-8. Epub 2018 Apr 23. Planta. 2018. PMID: 29687222
-
Recent advances in understanding female gametophyte development.F1000Res. 2018 Jun 20;7:F1000 Faculty Rev-804. doi: 10.12688/f1000research.14508.1. eCollection 2018. F1000Res. 2018. PMID: 29983913 Free PMC article. Review.
-
Focus on Flowering and Reproduction.Plant Physiol. 2017 Jan;173(1):1-4. doi: 10.1104/pp.16.01867. Plant Physiol. 2017. PMID: 28049854 Free PMC article. No abstract available.
-
In Silico Functional Prediction and Expression Analysis of C2H2 Zinc-Finger Family Transcription Factor Revealed Regulatory Role of ZmZFP126 in Maize Growth.Front Genet. 2021 Nov 5;12:770427. doi: 10.3389/fgene.2021.770427. eCollection 2021. Front Genet. 2021. PMID: 34804129 Free PMC article.
References
-
- Amedeo P, Habu Y, Afsar K, Mittelsten Scheid O, Paszkowski J (2000) Disruption of the plant gene MOM releases transcriptional silencing of methylated genes. Nature 405: 203–206 - PubMed
-
- Aravind L. (2000) The BED finger, a novel DNA-binding domain in chromatin-boundary-element-binding proteins and transposases. Trends Biochem Sci 25: 421–423 - PubMed
-
- Austin RS, Hiu S, Waese J, Ierullo M, Pasha A, Wang TT, Fan J, Foong C, Breit R, Desveaux D, Moses A, Provart NJ (2016) New BAR tools for mining expression data and exploring Cis-elements in Arabidopsis thaliana. Plant J 88: 490–504 - PubMed
-
- Bao Z, Zhang N, Hua J (2014) Endopolyploidization and flowering time are antagonistically regulated by checkpoint component MAD1 and immunity modulator MOS1. Nat Commun 5: 5628. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
