

Review
doi: 10.1038/nsmb.3324.
Genetic and epigenomic mechanisms of mammalian circadian transcription
Affiliations
- PMID: 27922611
- PMCID: PMC5497498
- DOI: 10.1038/nsmb.3324
Item in Clipboard
Review
Genetic and epigenomic mechanisms of mammalian circadian transcription
Nat Struct Mol Biol.
.
Abstract
The mammalian molecular clock comprises a complex network of transcriptional programs that integrates environmental signals with physiological pathways in a tissue-specific manner. Emerging technologies are extending knowledge of basic clock features by uncovering their underlying molecular mechanisms, thus setting the stage for a 'systems' view of the molecular clock. Here we discuss how recent data from genome-wide genetic and epigenetic studies have informed the understanding of clock function. In addition to its importance in human physiology and disease, the clock mechanism provides an ideal model to assess general principles of dynamic transcription regulation in vivo.
Figures
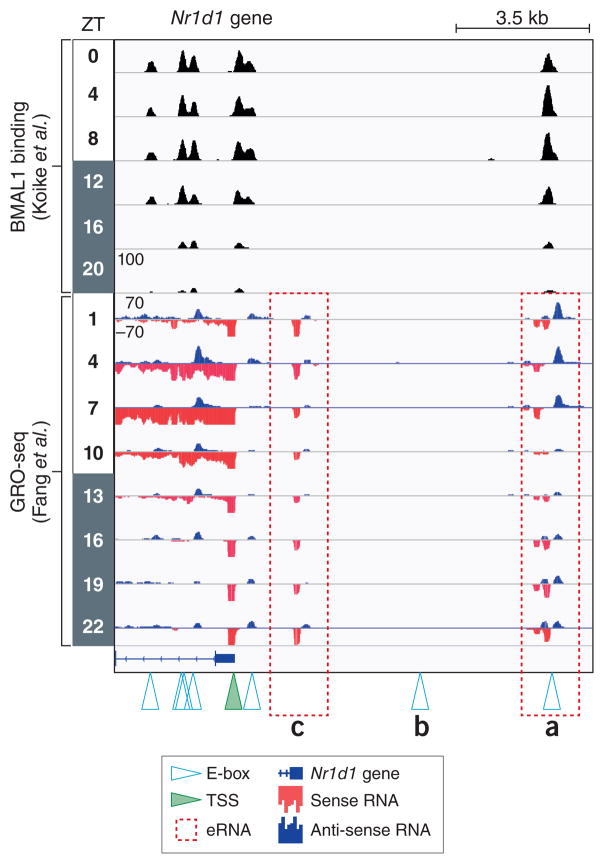
ChIP-seq data from Koike et al. juxtaposed to Global Run-On Sequencing (GRO-Seq) data from Fang et al. performed in mouse livers from circadian time points (ZT0 is light on, ZT 12 is lights off). Canonical CACGTG E-box motifs (denoted by open triangles) are bound by BMAL1 with the exception of (b). GRO-seq measures nascent transcription levels of genic and intergenic RNA, including short-lived enhancer RNA (eRNA). Two rhythmically expressed eRNAs (a) and (c) are present near the transcriptional start site (TSS, green triangle) of Nr1d1, but only one (a) is directly bound by BMAL1. Whether these eRNA are linked topologically or whether the eRNA in (c) is independent of BMAL1 activity is not known.
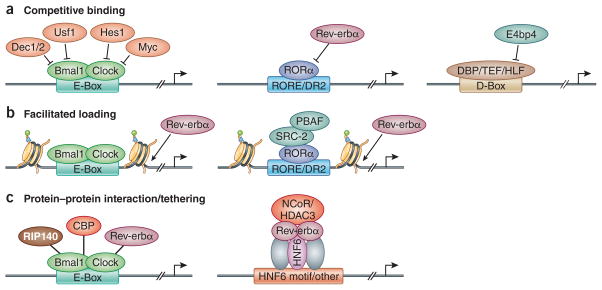
(a) Competitive binding among different TFs is observed at E-box, RORE, and D-box motifs. TF binding to the E-box motif, including DEC1/2, USF1, HES1, MYC can inhibit BMAL1 binding. Circadian expressed Rev-erbα competes with RORα, which results in oscillation of RORα binding. E4BP4 competes with other PAR-domain basic leucine zipper TFs DBP/TEF/HLF, which also leads to a robust diurnal switch on/off of their target gene transcription. (b) BMAL1 and CLOCK can function as pioneer TFs to promote the chromatin de-condensation during activation phase. Similarly, RORα could also recruit epigenetic modification machinery to remodel the chromatin structure to facilitate the binding of Rev-erbα. (c) PXDLS peptide motif, which is conserved in CBP, RIP140 and Rev-erbα, can interact with BMAL1/CLOCK and regulate circadian transcription. On the other hand, Rev-erbα can also be tethered to lineage determining TFs, such as HNF6 in the liver, to regulate different metabolic output processes in various tissues.
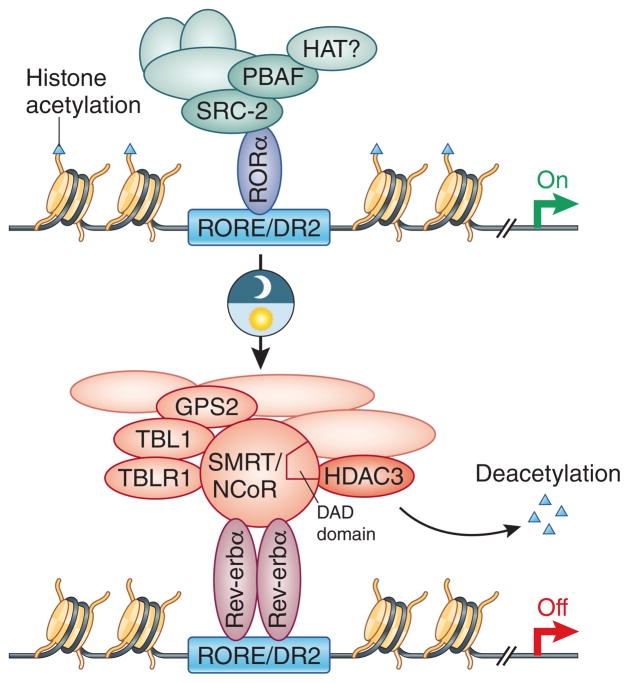
Rev-erb represses transcription by two known major mechanisms: 1) Competes with and jettisons RORα and associated co-activators from chromatin; and 2) Recruits N-CoR/SMRT corepressor complexes harboring HDAC3 to deacetylate nearby chromatin and heterochromatin. The deacetylase activation domain (DAD) of N-CoR and SMRT is required to stimulate HDAC3 activity.
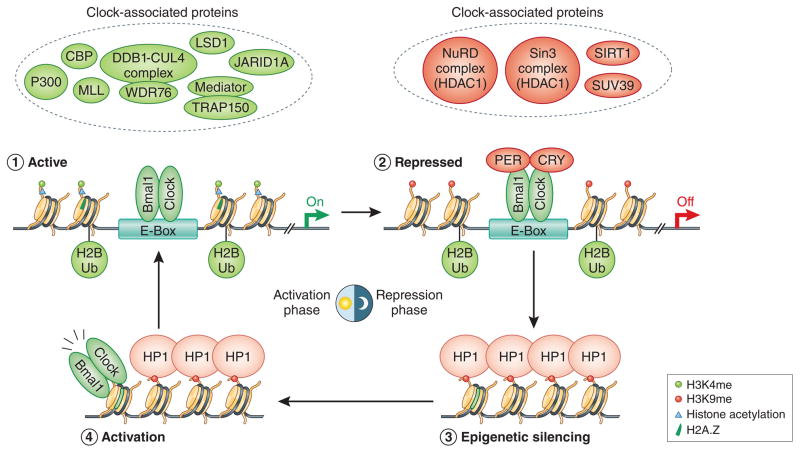
Recent work has started to unravel the temporal coordination of the molecular clock, including rhythmic changes to histone modifications, histone deposition and chromatin structure. The classical clock proteins (BMAL1/CLOCK/CRY/PER) recruit several chromatin-modifying proteins and complexes that control the epigenome.
References
-
- Bedrosian Ta, Fonken LK, Nelson RJ. Endocrine Effects of Circadian Disruption. Annu Rev Physiol. 2015;78:150724172241001. - PubMed
-
- Price JL, et al. double-time is a novel Drosophila clock gene that regulates period protein accumulation. Cell. 1998;94:83–95. - PubMed
-
- Hardin PE, Hall JC, Rosbash M. Feedback of the Drosophila period gene product on circadian cycling of its messenger RNA levels. Nature. 1990;343:536–540. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
