

Influenza A virus vaccines for swine
- PMID: 27923501
- PMCID: PMC8609643
- DOI: 10.1016/j.vetmic.2016.11.026
Influenza A virus vaccines for swine
Abstract
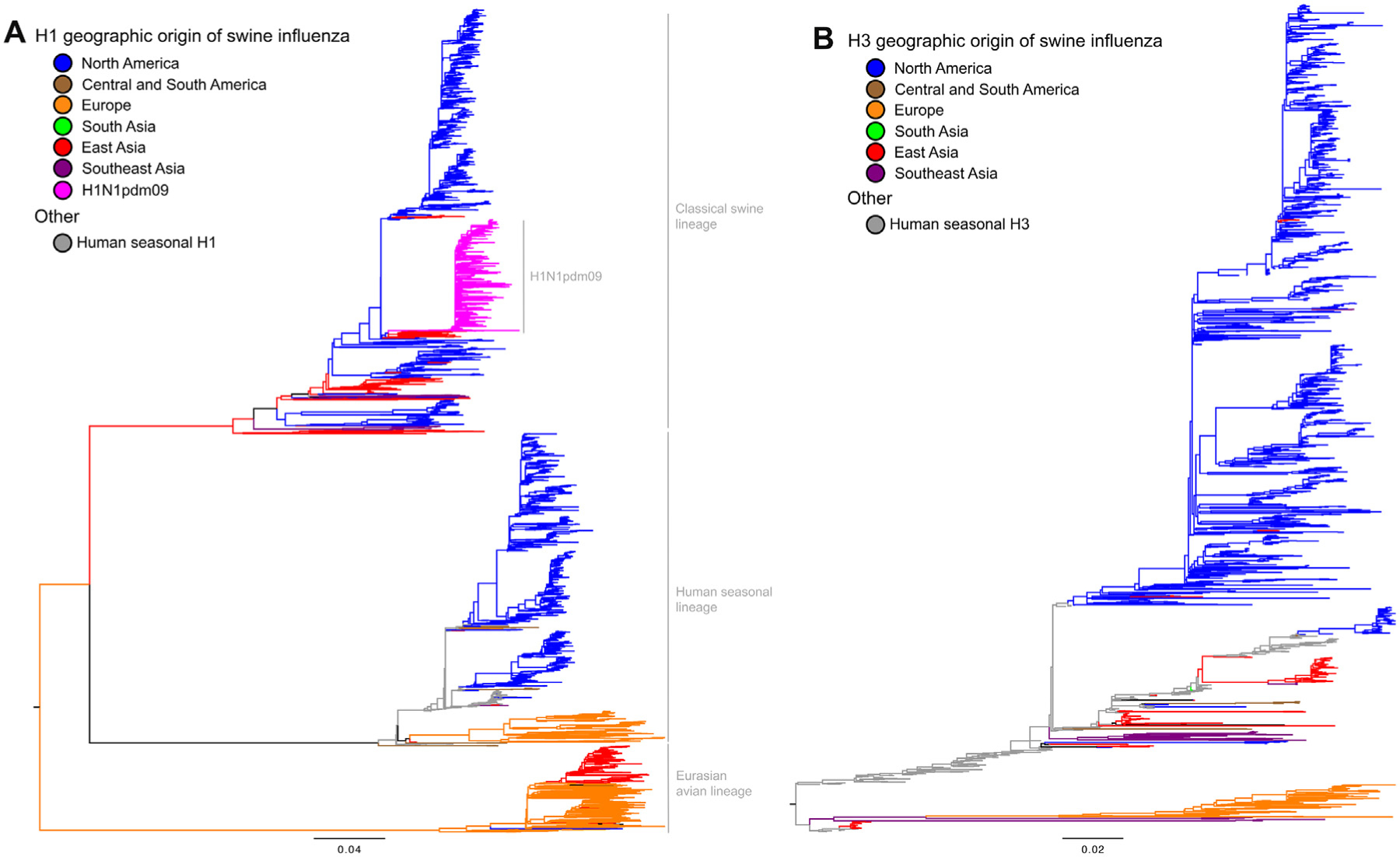
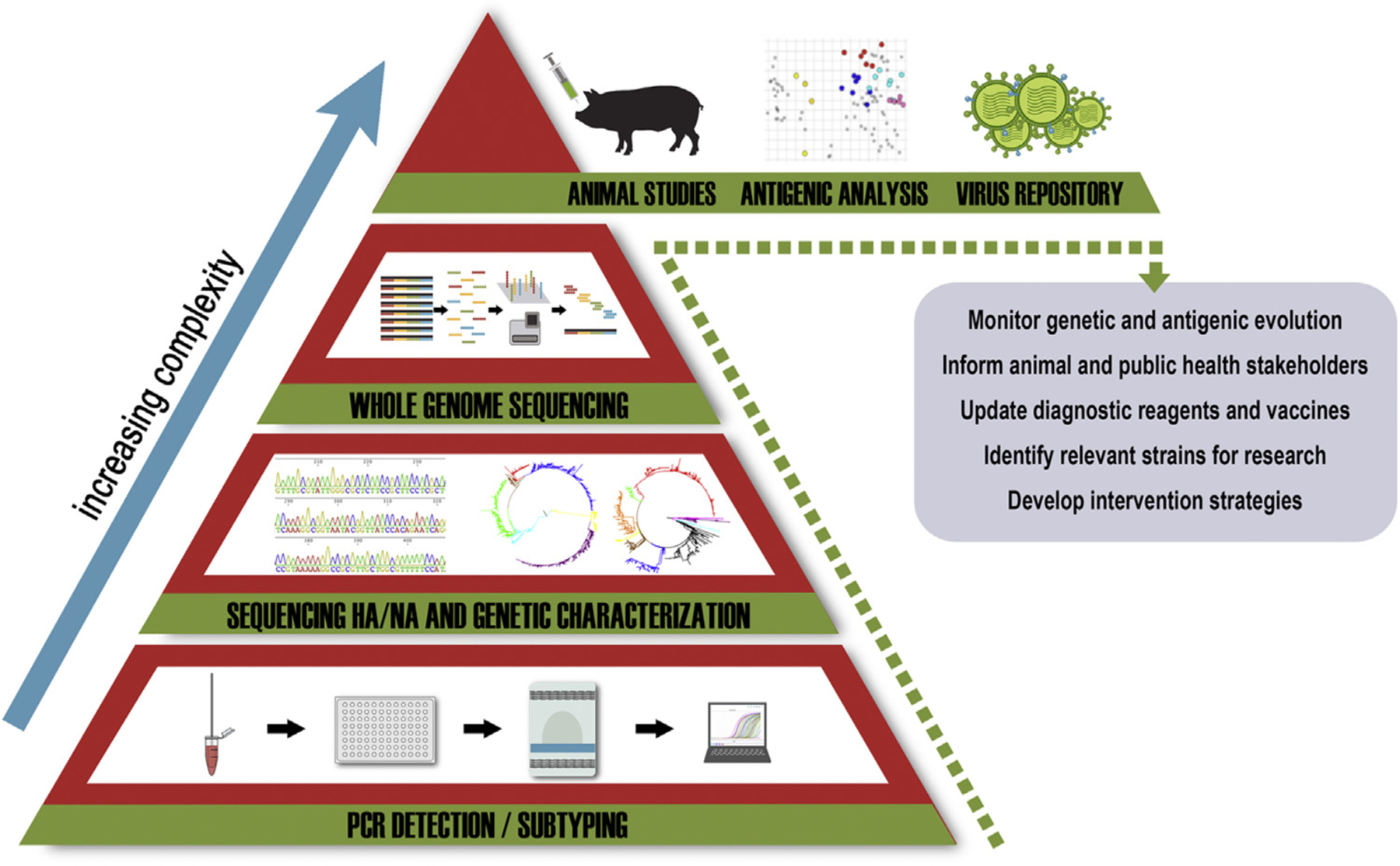
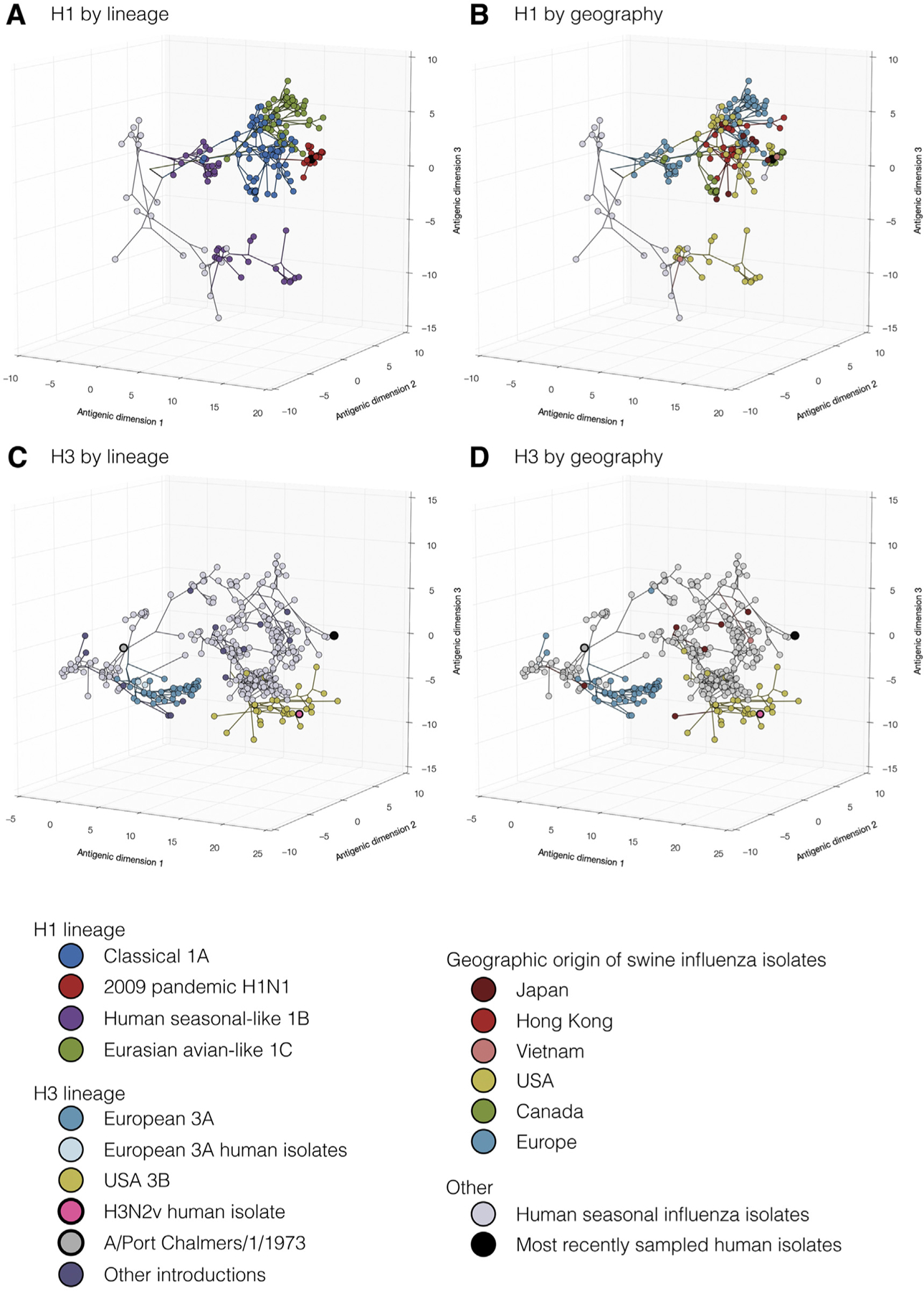
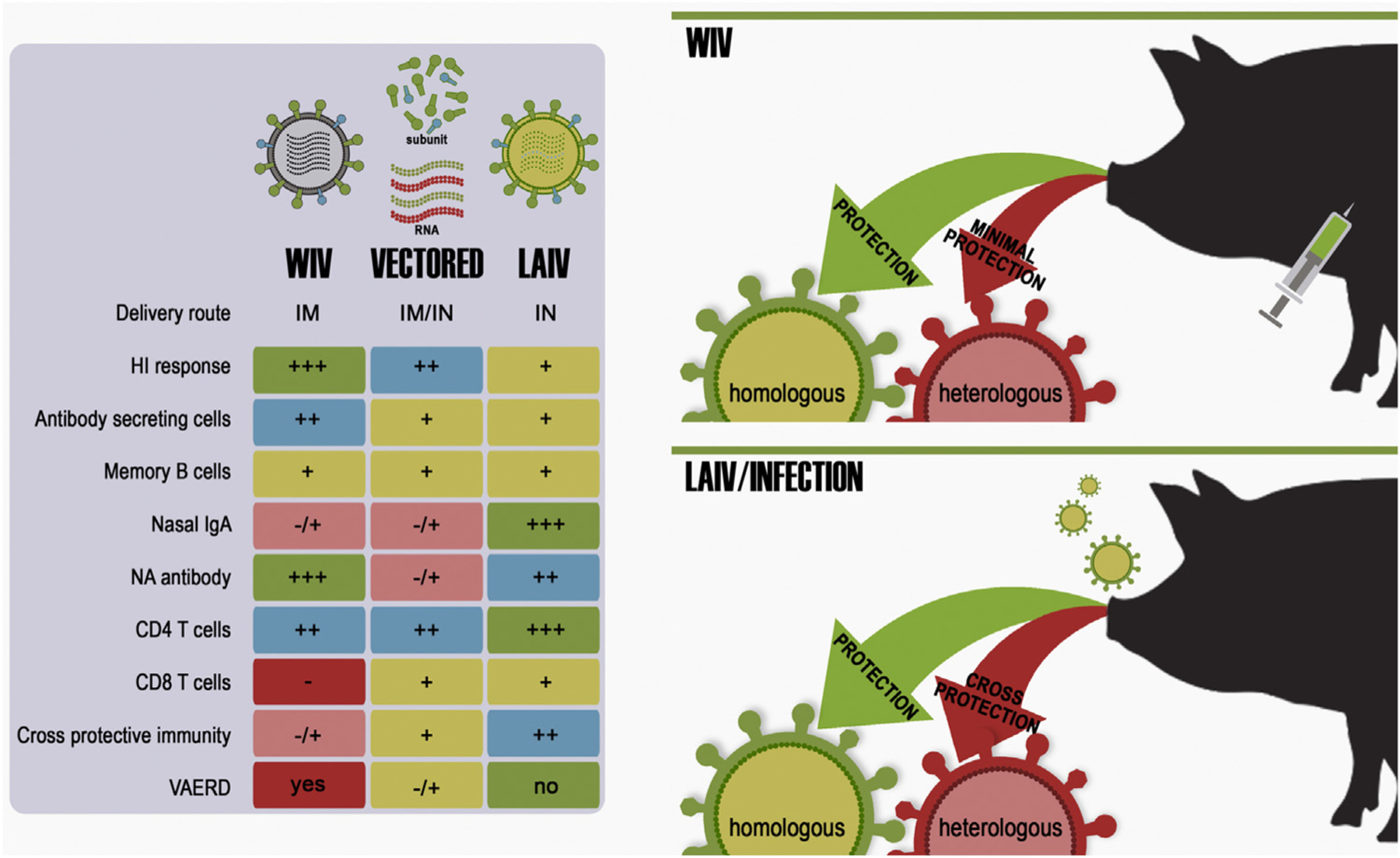
Economic losses due to influenza A virus (IAV) infections are substantial and a global problem, ranking among the top three major health challenges in the swine industry. Currently, H1 and H3 subtypes circulate in pigs globally associated with different combinations of N1 and N2 subtypes; however, the origin, gene constellation, and antigenic makeup of IAV vary greatly on different continents. Vaccination is one means of mitigating the effects of IAV disease, and vaccines are most effective if the strains included closely match the currently circulating strains in pigs. Genetic analyses provide panoramic views of the virus landscape at the sequence level and, thus, can aid in the selection of well-matched swine IAV vaccine strains, but is not sufficient alone. Additionally, a major challenge in selecting appropriate swine IAV vaccine strains is the co-circulation of multiple lineages of viruses in the same region, requiring multivalent or broadly cross-reacting antigens. Due to this complex IAV ecology in swine, new vaccination strategies and vaccine platforms are needed. The hemagglutinin (HA) viral protein is the major target of neutralizing antibodies, which are widely considered to be correlated with protection. Virus variants that are not recognized by previously elicited antibodies can render traditional vaccines that primarily elicit humoral responses ineffective, and therefore result in the need for vaccine strain reformulation and re-vaccination. In the future, new vaccine platforms may be on the market that will provide alternative options to those currently available. Nonetheless, a collaborative approach is needed to improve IAV vaccine strain selection for use in swine.
Keywords: H1N1; H1N2; H3N2; Influenza A virus; Swine; Vaccines.
Published by Elsevier B.V.
Conflict of interest statement
Conflicts of interest
The authors declare that there are no conflicts of interest.
Figures
References
-
- Allerson MW, Davies PR, Gramer MR, Torremorell M, 2014. Infection dynamics of pandemic 2009 H1N1 influenza virus in a two-site swine herd. Transbound. Emerg. Dis 61, 490–499. - PubMed
-
- Ampofo WK, Azziz-Baumgartner E, Bashir U, Cox NJ, Fasce R, Giovanni M, Grohmann G, Huang S, Katz J, Mironenko A, Mokhtari-Azad T, Sasono PM, Rahman M, Sawanpanyalert P, Siqueira M, Waddell AL, Waiboci L, Wood J, Zhang W, Ziegler T, Group WHOW, 2015. Strengthening the influenza vaccine virus selection and development process: report of the 3rd WHO informal consultation for improving influenza vaccine virus selection held at WHO headquarters, Geneva, Switzerland, 1–3 april 2014. Vaccine 33, 4368–4382. - PubMed
-
- Anderson TK, Campbell BA, Nelson MI, Lewis NS, Janas-Martindale A, Killian ML, Vincent AL, 2015. Characterization of co-circulating swine influenza A viruses in North America and the identification of a novel H1 genetic clade with antigenic significance. Virus Res. 201, 24–31. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
