

Estrogen activation of microglia underlies the sexually dimorphic differences in Nf1 optic glioma-induced retinal pathology
- PMID: 27923908
- PMCID: PMC5206494
- DOI: 10.1084/jem.20160447
Estrogen activation of microglia underlies the sexually dimorphic differences in Nf1 optic glioma-induced retinal pathology
Abstract
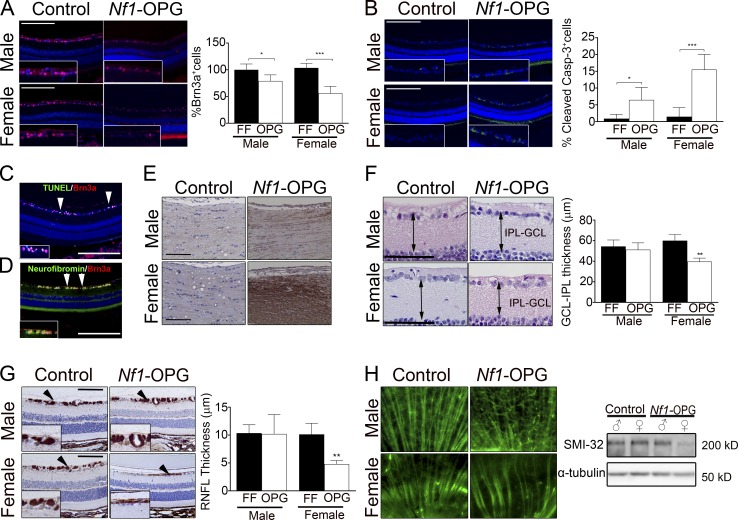
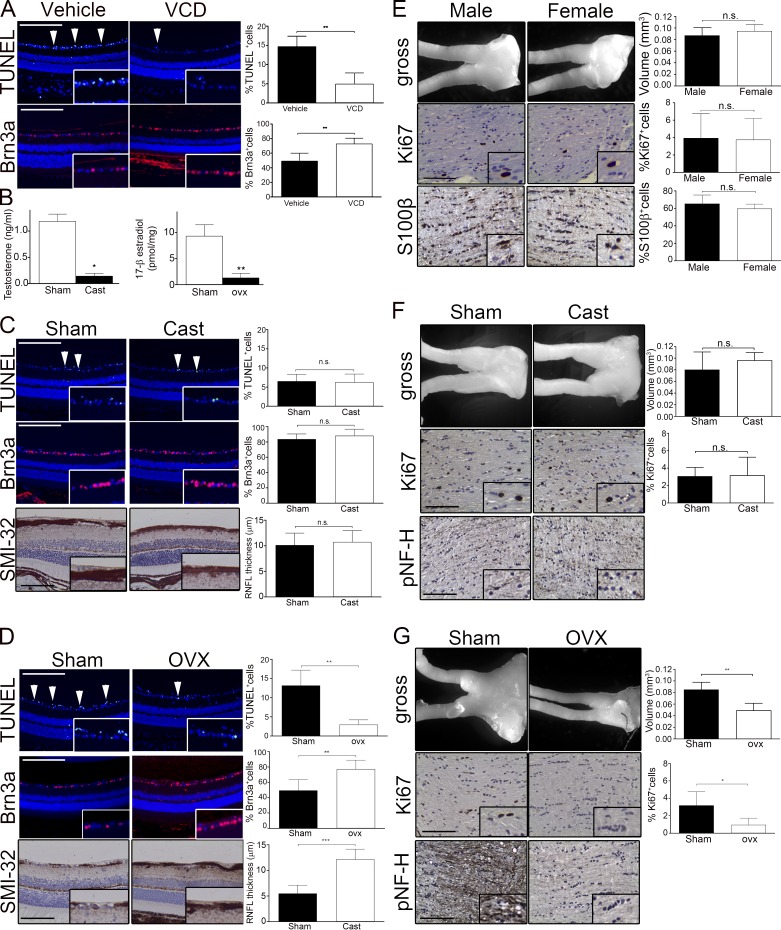
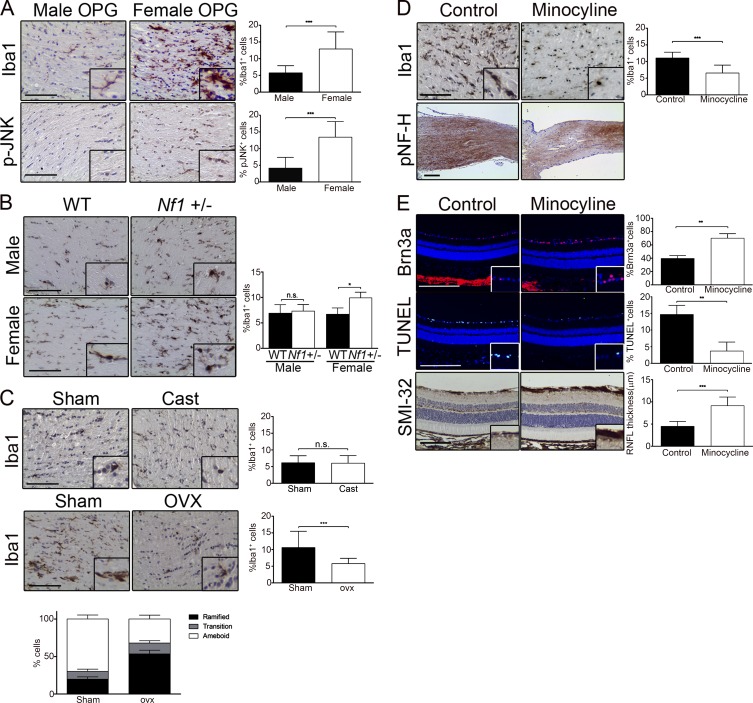
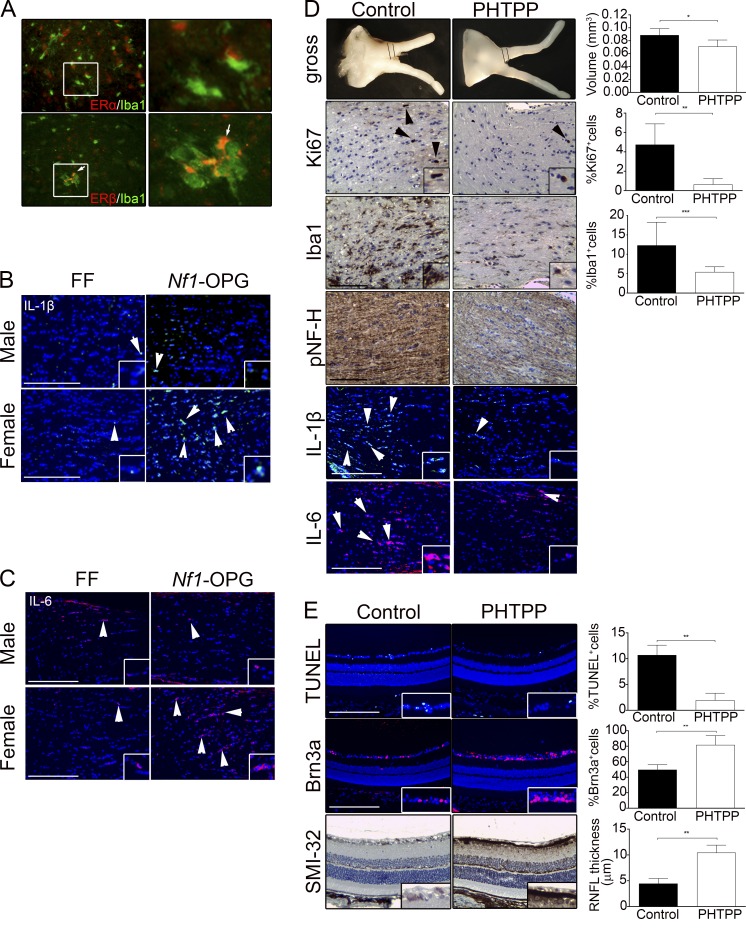
Children with neurofibromatosis type 1 (NF1) develop low-grade brain tumors throughout the optic pathway. Nearly 50% of children with optic pathway gliomas (OPGs) experience visual impairment, and few regain their vision after chemotherapy. Recent studies have revealed that girls with optic nerve gliomas are five times more likely to lose vision and require treatment than boys. To determine the mechanism underlying this sexually dimorphic difference in clinical outcome, we leveraged Nf1 optic glioma (Nf1-OPG) mice. We demonstrate that female Nf1-OPG mice exhibit greater retinal ganglion cell (RGC) loss and only females have retinal nerve fiber layer (RNFL) thinning, despite mice of both sexes harboring tumors of identical volumes and proliferation. Female gonadal sex hormones are responsible for this sexual dimorphism, as ovariectomy, but not castration, of Nf1-OPG mice normalizes RGC survival and RNFL thickness. In addition, female Nf1-OPG mice have threefold more microglia than their male counterparts, and minocycline inhibition of microglia corrects the retinal pathology. Moreover, pharmacologic inhibition of microglial estrogen receptor-β (ERβ) function corrects the retinal abnormalities in female Nf1-OPG mice. Collectively, these studies establish that female gonadal sex hormones underlie the sexual dimorphic differences in Nf1 optic glioma-induced retinal dysfunction by operating at the level of tumor-associated microglial activation.
© 2017 Toonen et al.
Figures
Similar articles
-
Defining the temporal course of murine neurofibromatosis-1 optic gliomagenesis reveals a therapeutic window to attenuate retinal dysfunction.Neuro Oncol. 2017 Jun 1;19(6):808-819. doi: 10.1093/neuonc/now267. Neuro Oncol. 2017. PMID: 28039362 Free PMC article.
-
Measurements of Retinal Nerve Fiber Thickness and Ganglion Cell Complex in Neurofibromatosis Type 1, with and Without Optic Pathway Gliomas: A Case Series.Curr Eye Res. 2018 Mar;43(3):424-427. doi: 10.1080/02713683.2017.1408130. Epub 2017 Nov 29. Curr Eye Res. 2018. PMID: 29185816
-
Estrogen-induced glial IL-1β mediates extrinsic retinal ganglion cell vulnerability in murine Nf1 optic glioma.Ann Clin Transl Neurol. 2024 Mar;11(3):812-818. doi: 10.1002/acn3.51995. Epub 2024 Jan 16. Ann Clin Transl Neurol. 2024. PMID: 38229454 Free PMC article.
-
Insights into optic pathway glioma vision loss from mouse models of neurofibromatosis type 1.J Neurosci Res. 2019 Jan;97(1):45-56. doi: 10.1002/jnr.24250. Epub 2018 Apr 28. J Neurosci Res. 2019. PMID: 29704429 Free PMC article. Review.
-
Optic Pathway Gliomas in Neurofibromatosis Type 1.J Child Neurol. 2018 Jan;33(1):73-81. doi: 10.1177/0883073817739509. J Child Neurol. 2018. PMID: 29246098 Free PMC article. Review.
Cited by
-
JAM-A functions as a female microglial tumor suppressor in glioblastoma.Neuro Oncol. 2020 Nov 26;22(11):1591-1601. doi: 10.1093/neuonc/noaa148. Neuro Oncol. 2020. PMID: 32592484 Free PMC article.
-
Immunohistochemical Markers for Prospective Studies in Neurofibromatosis-1 Porcine Models.J Histochem Cytochem. 2017 Oct;65(10):607-618. doi: 10.1369/0022155417729357. Epub 2017 Aug 28. J Histochem Cytochem. 2017. PMID: 28846462 Free PMC article.
-
Trends in the global burden of vision loss among the older adults from 1990 to 2019.Front Public Health. 2024 Apr 4;12:1324141. doi: 10.3389/fpubh.2024.1324141. eCollection 2024. Front Public Health. 2024. PMID: 38638474 Free PMC article.
-
Estrogen Interactions With Lipid Rafts Related to Neuroprotection. Impact of Brain Ageing and Menopause.Front Neurosci. 2018 Mar 6;12:128. doi: 10.3389/fnins.2018.00128. eCollection 2018. Front Neurosci. 2018. PMID: 29559883 Free PMC article. Review.
-
Implications of new understandings of gliomas in children and adults with NF1: report of a consensus conference.Neuro Oncol. 2020 Jun 9;22(6):773-784. doi: 10.1093/neuonc/noaa036. Neuro Oncol. 2020. PMID: 32055852 Free PMC article.
References
-
- Azevedo E.P., Ledo J.H., Barbosa G., Sobrinho M., Diniz L., Fonseca A.C., Gomes F., Romão L., Lima F.R., Palhano F.L., et al. . 2013. Activated microglia mediate synapse loss and short-term memory deficits in a mouse model of transthyretin-related oculoleptomeningeal amyloidosis. Cell Death Dis. 4:e789 10.1038/cddis.2013.325 - DOI - PMC - PubMed
-
- Bajenaru M.L., Hernandez M.R., Perry A., Zhu Y., Parada L.F., Garbow J.R., and Gutmann D.H.. 2003. Optic nerve glioma in mice requires astrocyte Nf1 gene inactivation and Nf1 brain heterozygosity. Cancer Res. 63:8573–8577. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
