

Regulation of Prp43-mediated disassembly of spliceosomes by its cofactors Ntr1 and Ntr2
- PMID: 27923990
- PMCID: PMC5397206
- DOI: 10.1093/nar/gkw1225
Regulation of Prp43-mediated disassembly of spliceosomes by its cofactors Ntr1 and Ntr2
Abstract
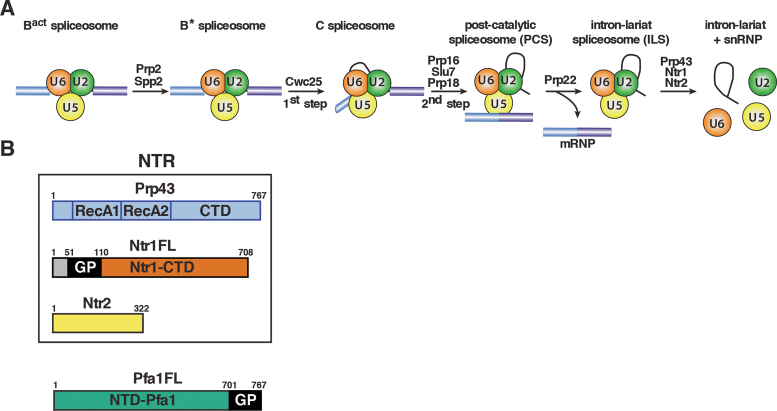
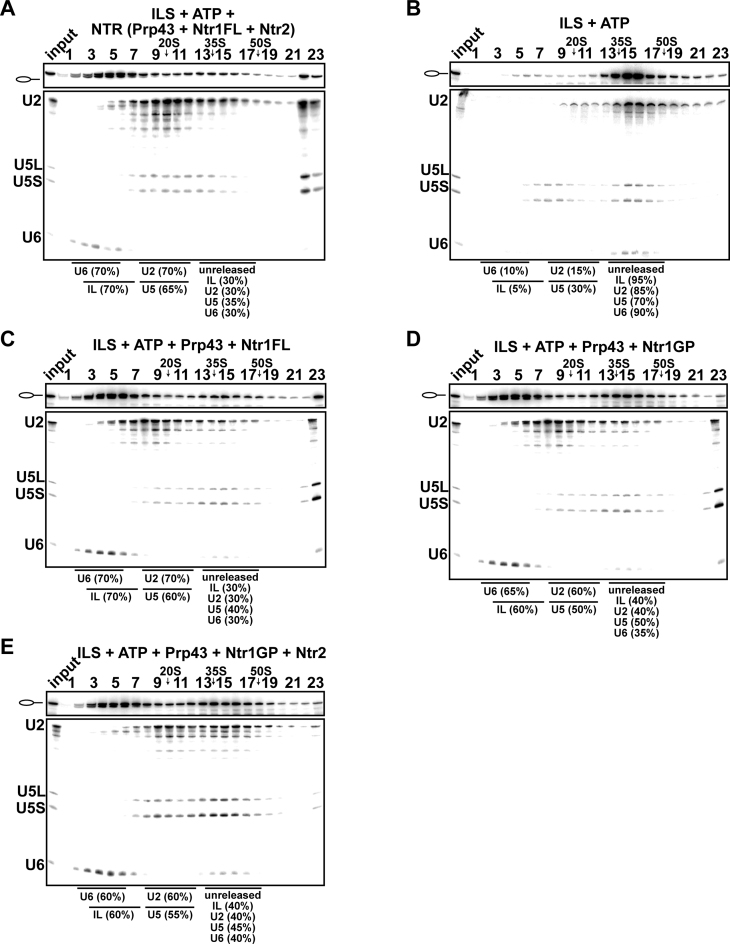
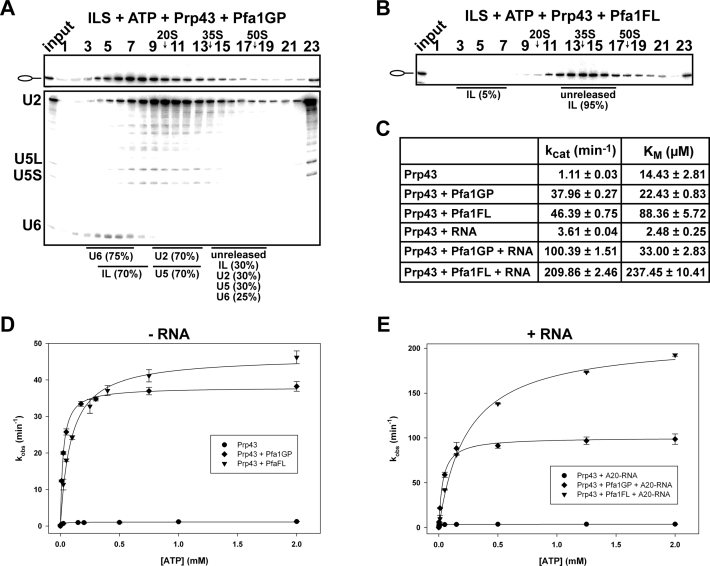
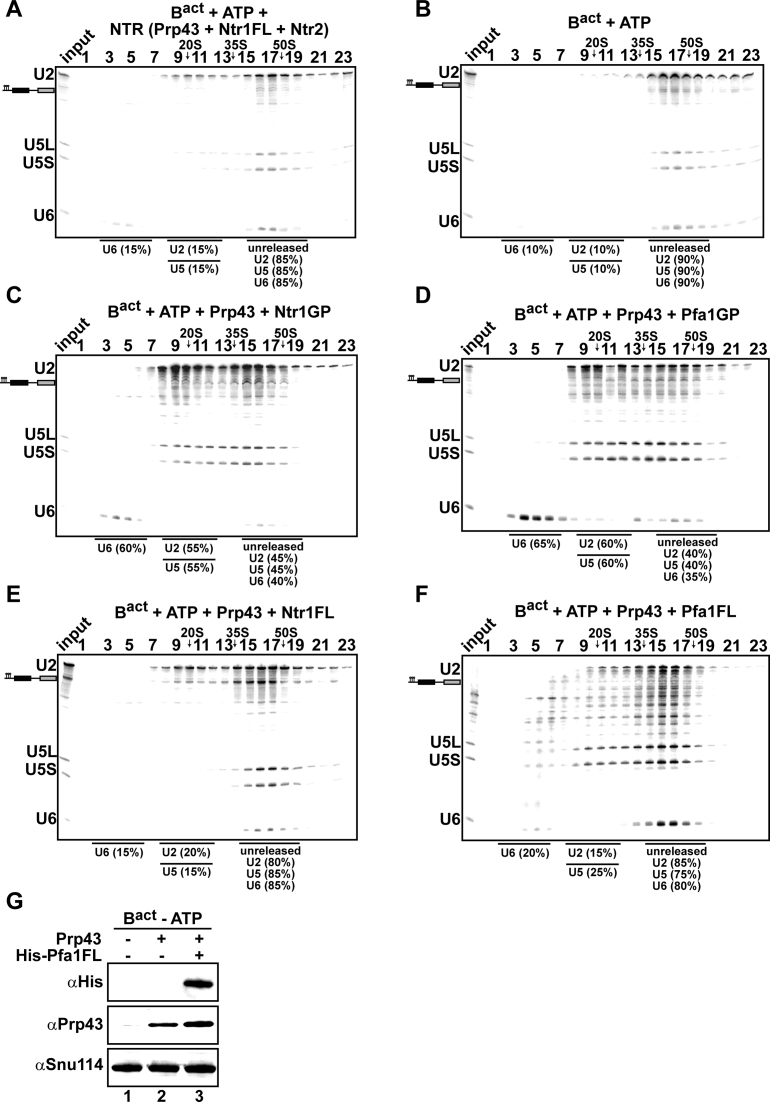
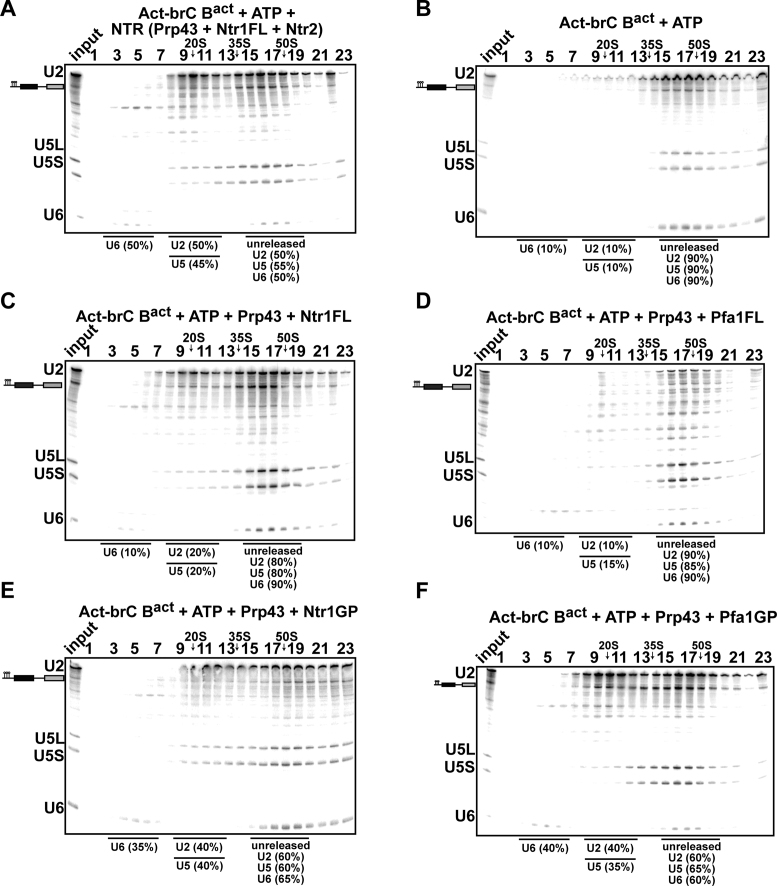
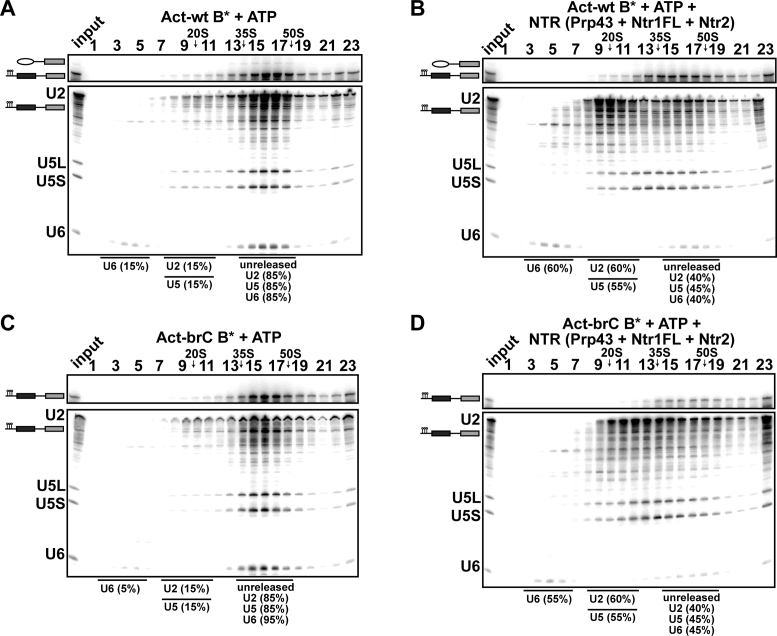
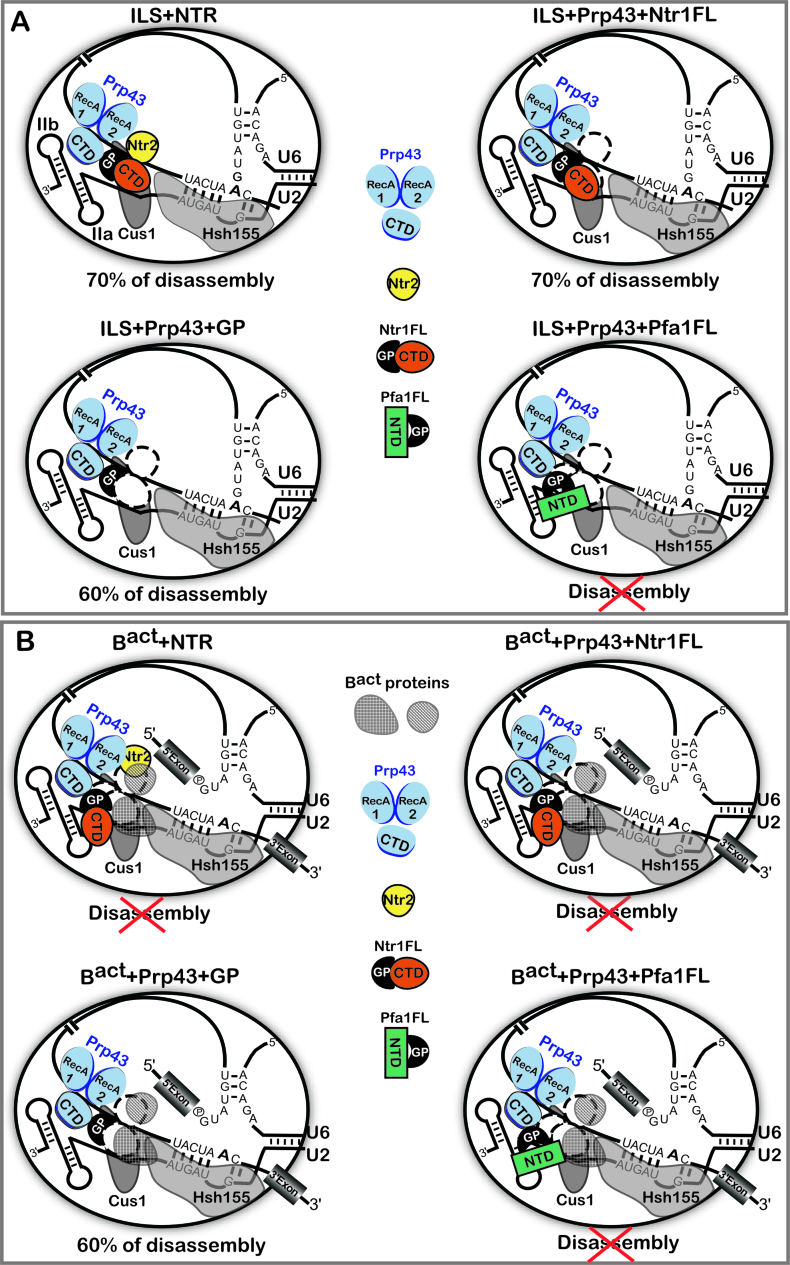
The DEAH-box NTPase Prp43 disassembles spliceosomes in co-operation with the cofactors Ntr1/Spp382 and Ntr2, forming the NTR complex. How Prp43 is regulated by its cofactors to discard selectively only intron-lariat spliceosomes (ILS) and defective spliceosomes and to prevent disassembly of earlier and properly assembled/wild-type spliceosomes remains unclear. First, we show that Ntr1΄s G-patch motif (Ntr1GP) can be replaced by the GP motif of Pfa1/Sqs1, a Prp43΄s cofactor in ribosome biogenesis, demonstrating that the specific function of Ntr1GP is to activate Prp43 for spliceosome disassembly and not to guide Prp43 to its binding site in the spliceosome. Furthermore, we show that Ntr1΄s C-terminal domain (CTD) plays a safeguarding role by preventing Prp43 from disrupting wild-type spliceosomes other than the ILS. Ntr1 and Ntr2 can also discriminate between wild-type and defective spliceosomes. In both type of spliceosomes, Ntr1-CTD impedes Prp43-mediated disassembly while the Ntr1GP promotes disassembly. Intriguingly, Ntr2 plays a specific role in defective spliceosomes, likely by stabilizing Ntr1 and allowing Prp43 to enter a productive interaction with the GP motif of Ntr1. Our data indicate that Ntr1 and Ntr2 act as 'doorkeepers' and suggest that both cofactors inspect the RNP structure of spliceosomal complexes thereby targeting suboptimal spliceosomes for Prp43-mediated disassembly.
© The Author(s) 2016. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
References
-
- Wahl M.C., Will C.L., Lührmann R.. The spliceosome: design principles of a dynamic RNP machine. Cell. 2009; 136:701–718. - PubMed
-
- Cordin O., Hahn D., Beggs J.D.. Structure, function and regulation of spliceosomal RNA helicases. Curr. Opin. Cell Biol. 2012; 24:431–438. - PubMed
-
- O’Day C.L., Dalbadie-McFarland G., Abelson J.. The Saccharomyces cerevisiae Prp5 protein has RNA-dependent ATPase activity with specificity for U2 small nuclear RNA. J. Biol. Chem. 1996; 271:33261–33267. - PubMed
-
- Gozani O., Feld R., Reed R.. Evidence that sequence-independent binding of highly conserved U2 snRNP proteins upstream of the branch site is required for assembly of spliceosomal complex A. Genes Dev. 1996; 10:233–243. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
