

Mdm2 as a chromatin modifier
- PMID: 27927750
- PMCID: PMC5439376
- DOI: 10.1093/jmcb/mjw046
Mdm2 as a chromatin modifier
Abstract
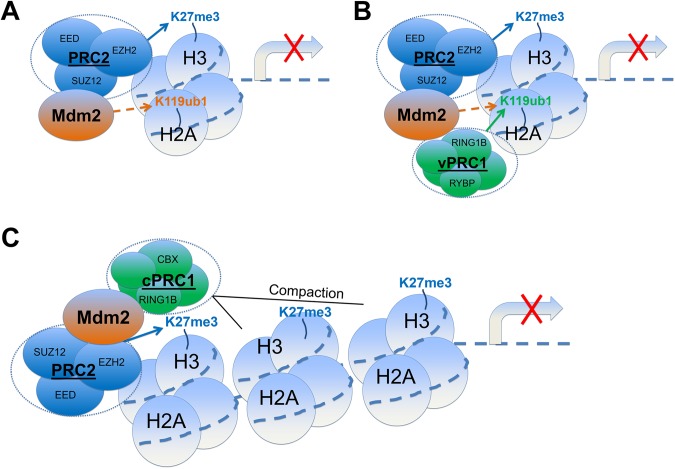
Mdm2 is the key negative regulator of the tumour suppressor p53, making it an attractive target for anti-cancer drug design. We recently identified a new role of Mdm2 in gene repression through its direct interaction with several proteins of the polycomb group (PcG) family. PcG proteins form polycomb repressive complexes PRC1 and PRC2. PRC2 (via EZH2) mediates histone 3 lysine 27 (H3K27) trimethylation, and PRC1 (via RING1B) mediates histone 2A lysine 119 (H2AK119) monoubiquitination. Both PRCs mostly support a compact and transcriptionally silent chromatin structure. We found that Mdm2 regulates a gene expression profile similar to that of PRC2 independent of p53. Moreover, Mdm2 promotes the stemness of murine induced pluripotent stem cells and human mesenchymal stem cells, and supports the survival of tumour cells. Mdm2 is recruited to target gene promoters by the PRC2 member and histone methyltransferase EZH2, and enhances PRC-dependent repressive chromatin modifications, specifically H3K27me3 and H2AK119ub1. Mdm2 also cooperates in gene repression with the PRC1 protein RING1B, a H2AK119 ubiquitin ligase. Here we discuss the possible implications of these p53-independent functions of Mdm2 in chromatin dynamics and in the stem cell phenotype. We propose that the p53-independent functions of Mdm2 should be taken into account for cancer drug design. So far, the majority of clinically tested Mdm2 inhibitors target its binding to p53 but do not affect the new functions of Mdm2 described here. However, when targeting the E3 ligase activity of Mdm2, a broader spectrum of its oncogenic activities might become druggable.
Keywords: EZH2; Mdm2; histone methylation; histone ubiquitination; polycomb repressor complex.
© The Author (2016). Published by Oxford University Press on behalf of Journal of Molecular Cell Biology, IBCB, SIBS, CAS.
Figures
References
-
- Alt J.R., Bouska A., Fernandez M.R., et al. (2005). Mdm2 binds to Nbs1 at sites of DNA damage and regulates double strand break repair. J. Biol. Chem. 280, 18771–18781. - PubMed
-
- Armstrong J.F., Kaufman M.H., Harrison D.J., et al. (1995). High-frequency developmental abnormalities in p53-deficient mice. Curr. Biol. 5, 931–936. - PubMed
-
- Arva N.C., Gopen T.R., Talbott K.E., et al. (2005). A chromatin-associated and transcriptionally inactive p53-Mdm2 complex occurs in mdm2 SNP309 homozygous cells. J. Biol. Chem. 280, 26776–26787. - PubMed
-
- Ben-Saadon R., Zaaroor D., Ziv T., et al. (2006). The polycomb protein Ring1B generates self atypical mixed ubiquitin chains required for its in vitro histone H2A ligase activity. Mol. Cell 24, 701–711. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
