

RNA processing body (P-body) dynamics in mesophyll protoplasts re-initiating cell division
- PMID: 27928633
- PMCID: PMC5487831
- DOI: 10.1007/s00709-016-1053-0
RNA processing body (P-body) dynamics in mesophyll protoplasts re-initiating cell division
Abstract
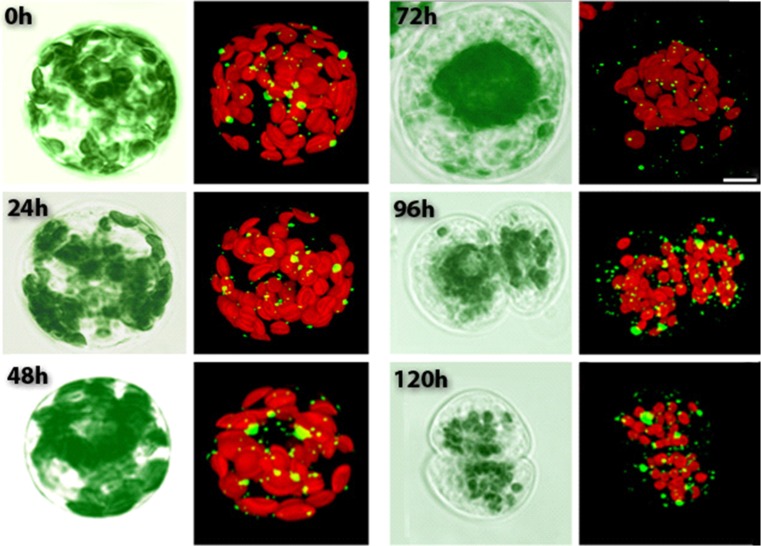
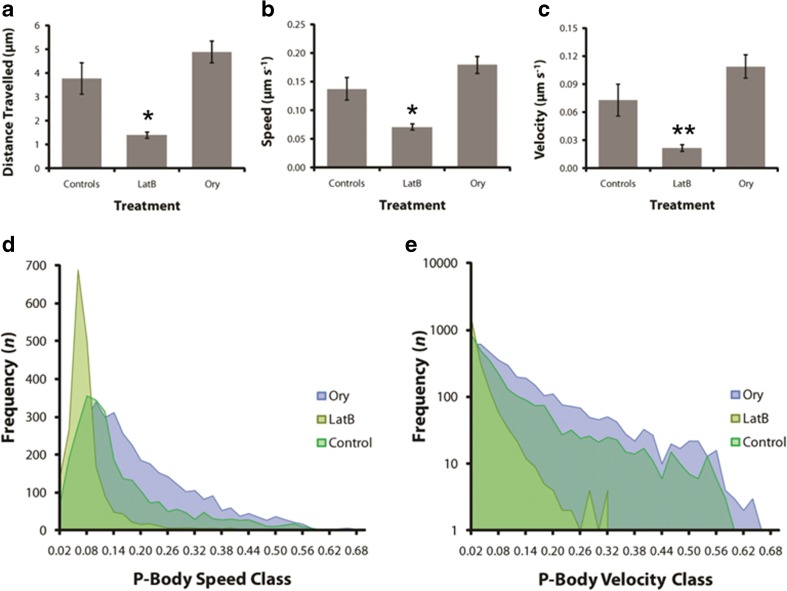
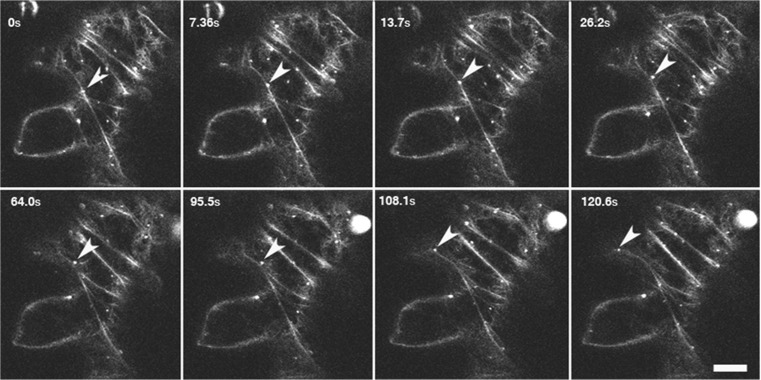
The ability of plants to regenerate lies in the capacity of differentiated cells to reprogram and re-enter the cell cycle. Reprogramming of cells requires changes in chromatin organisation and gene expression. However, there has been less focus on changes at the post transcription level. We have investigated P-bodies, sites of post transcriptional gene regulation, in plant cell reprogramming in cultured mesophyll protoplasts; by using a YFP-VARICOSE (YFP-VCSc) translational fusion. We showed an early increase in P-body number and volume, followed by a decline, then a subsequent continued increase in P-body number and volume as cell division was initiated and cell proliferation continued. We infer that plant P-bodies have a role to play in reprogramming the mature cell and re-initiating the cell division cycle. The timing of the first phase is consistent with the degredation of messages no longer required, as the cell transits to the division state, and may also be linked to the stress response associated with division induction in cultured cells. The subsequent increase in P-body formation, with partitioning to the daughter cells during the division process, suggests a role in the cell cycle and its re-initiation in daughter cells. P-bodies were shown to be mobile in the cytoplasm and show actin-based motility which facilitates their post-transcriptional role and partitioning to daughter cells.
Keywords: Dedifferentiation; P-bodies; Protoplast division; RNA decapping; RNA degredation; RNA storage.
Conflict of interest statement
The authors have no conflict of interest.
Figures
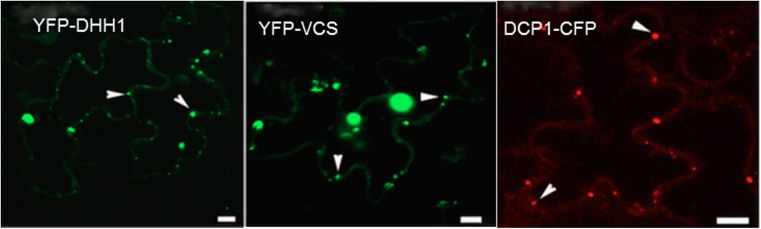
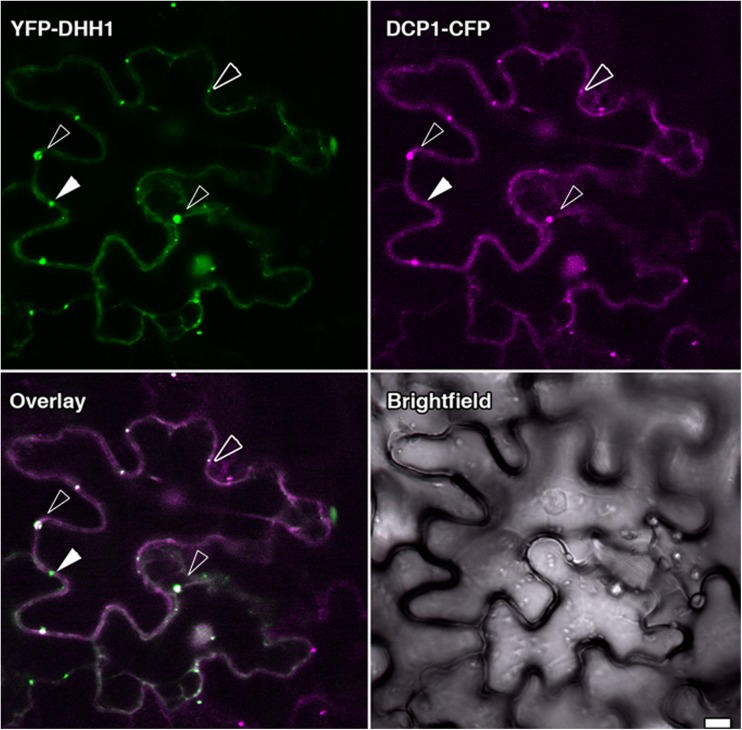
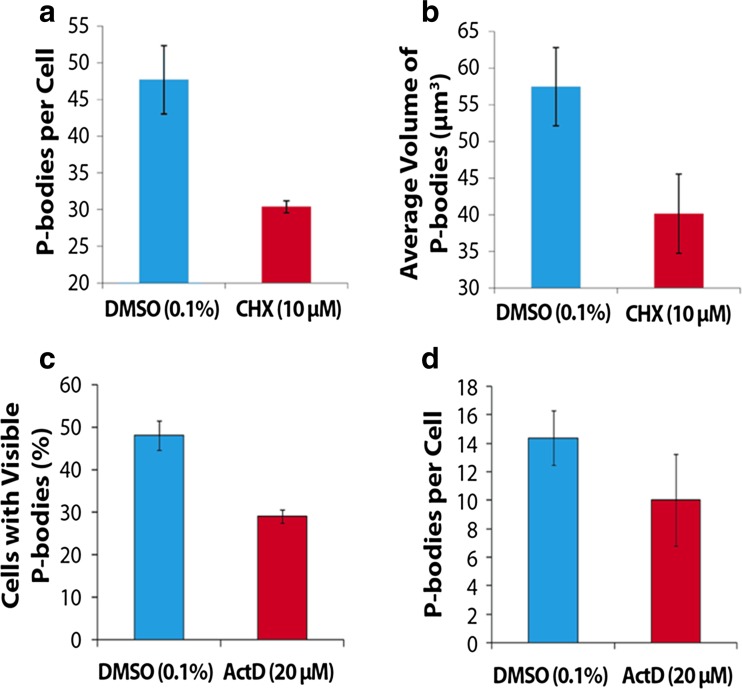
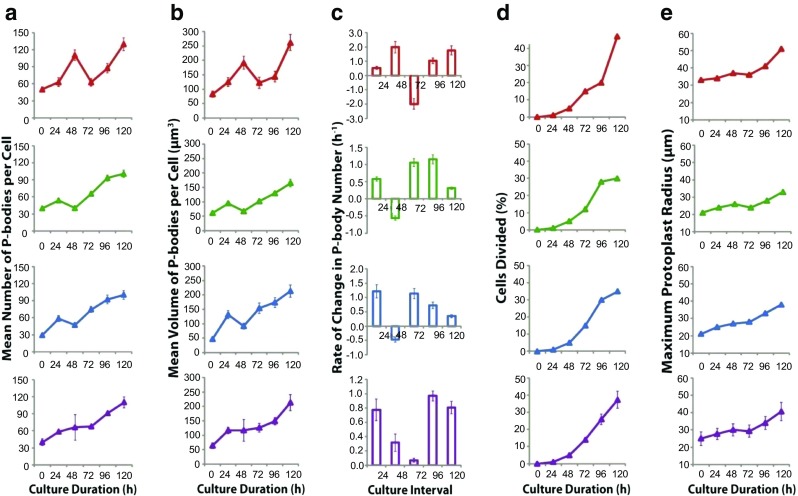

References
-
- Abràmoff MD, Magalhães PJ, Ram SJ. Image processing with ImageJ. Biophotonics Intern. 2004;11:36–42.
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
