

Deficiency in the Ubiquitin Conjugating Enzyme UBE2A in Alzheimer's Disease (AD) is Linked to Deficits in a Natural Circular miRNA-7 Sponge (circRNA; ciRS-7)
- PMID: 27929395
- PMCID: PMC5192492
- DOI: 10.3390/genes7120116
Deficiency in the Ubiquitin Conjugating Enzyme UBE2A in Alzheimer's Disease (AD) is Linked to Deficits in a Natural Circular miRNA-7 Sponge (circRNA; ciRS-7)
Abstract
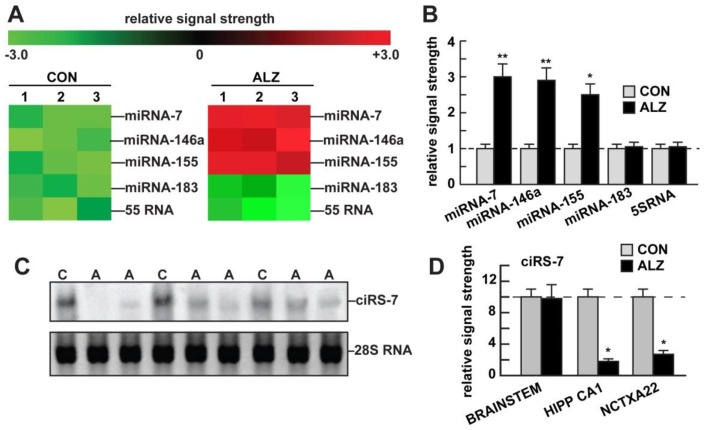
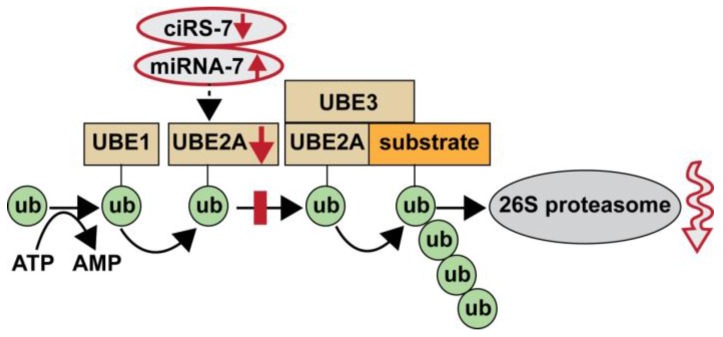
Our understanding of the highly specialized functions for small non-coding single-stranded RNA (ssRNA) in the transcriptome of the human central nervous system (CNS) continues to evolve. Circular RNAs (circRNAs), a recently discovered class of ssRNA enriched in the brain and retina, are extremely stable and intrinsically resilient to degradation by exonuclease. Conventional methods of ssRNA, microRNA (miRNA), or messenger RNA (mRNA) detection and quantitation requiring free ribonucleotide ends may have considerably underestimated the quantity and significance of CNS circRNA in the CNS. Highly-specific small ssRNAs, such as the ~23 nucleotide (nt) Homo sapien microRNA-7 (hsa-miRNA-7; chr 9q21.32), are not only abundant in the human limbic system but are, in addition, associated with a ~1400 nt circRNA for miRNA-7 (ciRS-7) in the same anatomical region. Structurally, ciRS-7 contains about ~70 tandem anti-miRNA-7 sequences and acts as an endogenous, anti-complementary miRNA-7 "sponge" that attracts, binds, and, hence, quenches, natural miRNA-7 functions. Using a combination of DNA and miRNA array technologies, enhanced LED-Northern and Western blot hybridization, and the magnesium-dependent exoribonuclease and circRNA-sensitive probe RNaseR, here we provide evidence of a significantly misregulated ciRS-7-miRNA-7-UBE2A circuit in sporadic Alzheimer's disease (AD) neocortex (Brodmann A22) and hippocampal CA1. Deficits in ciRS-7-mediated "sponging events", resulting in excess ambient miRNA-7 appear to drive the selective down-regulation in the expression of miRNA-7-sensitive mRNA targets, such as that encoding the ubiquitin conjugating enzyme E2A (UBE2A; chr Xq24). UBE2A, which normally serves as a central effector in the ubiquitin-26S proteasome system, coordinates the clearance of amyloid peptides via proteolysis, is known to be depleted in sporadic AD brain and, hence, contributes to amyloid accumulation and the formation of senile plaque deposits. Dysfunction of circRNA-miRNA-mRNA regulatory systems appears to represent another important layer of epigenetic control over pathogenic gene expression programs in the human CNS that are targeted by the sporadic AD process.
Keywords: Alzheimer’s disease (AD); circular RNA (circRNA); genetic control; miRNA-7; proteasome; proteolysis; ubiquitin conjugating enzyme E2A (UBE2A).
Conflict of interest statement
The authors declare no conflict of interest. The founding sponsors had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, and in the decision to publish the results.
Figures

References
-
- Ribonuclease R, E. coli. [(accessed on 17 November 2016)]. Available online: http://www.epibio.com/docs/default-source/protocols/ribonuclease-r-e-col....
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
