

A mechanism for acetylcholine receptor gating based on structure, coupling, phi, and flip
- PMID: 27932572
- PMCID: PMC5217088
- DOI: 10.1085/jgp.201611673
A mechanism for acetylcholine receptor gating based on structure, coupling, phi, and flip
Abstract
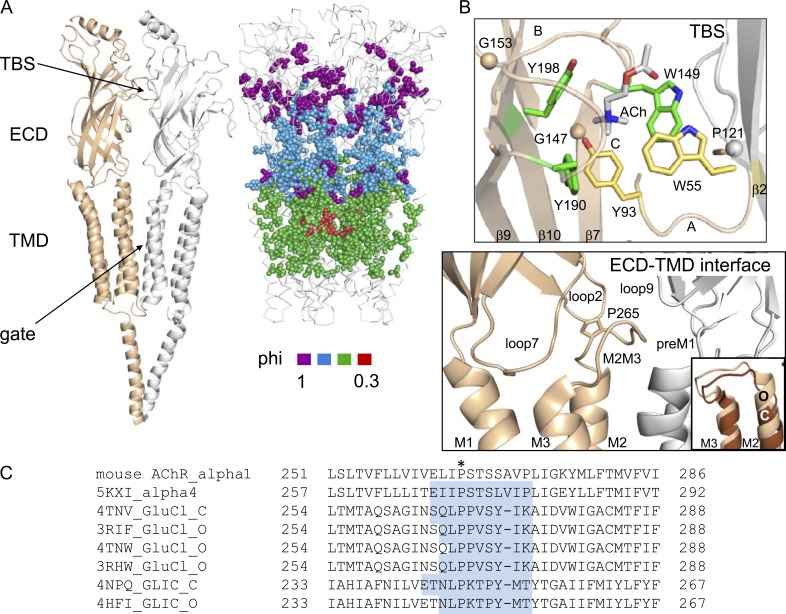
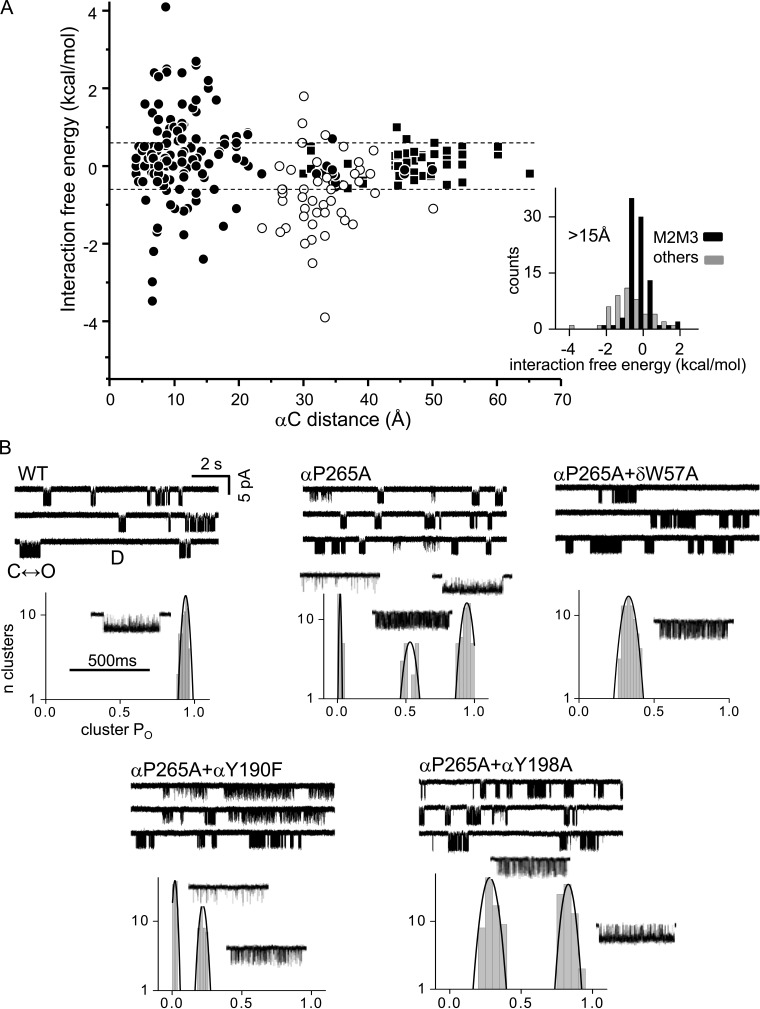
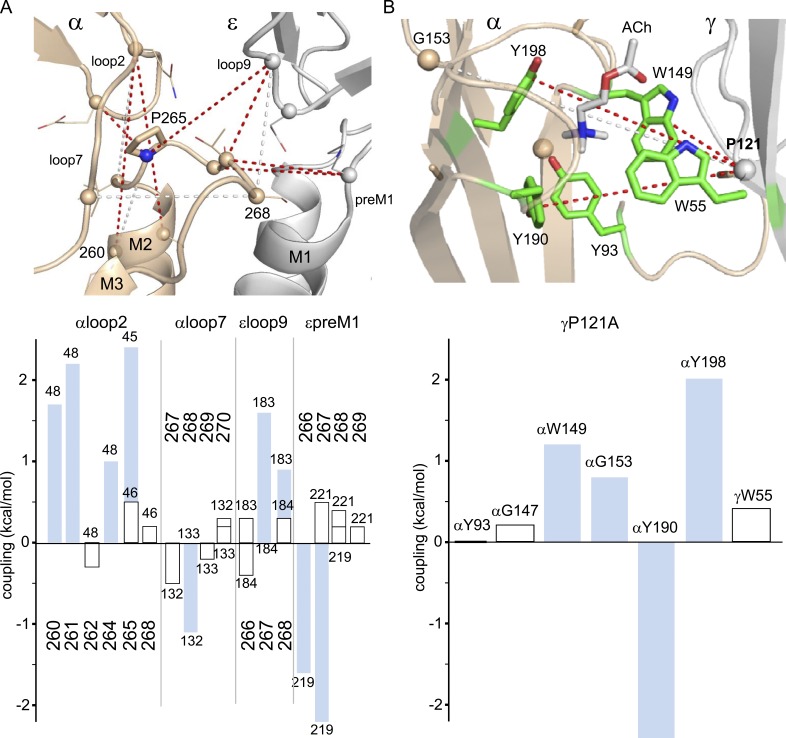
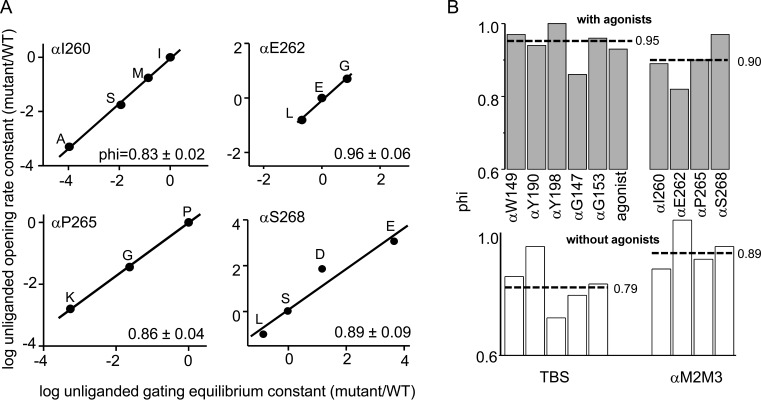
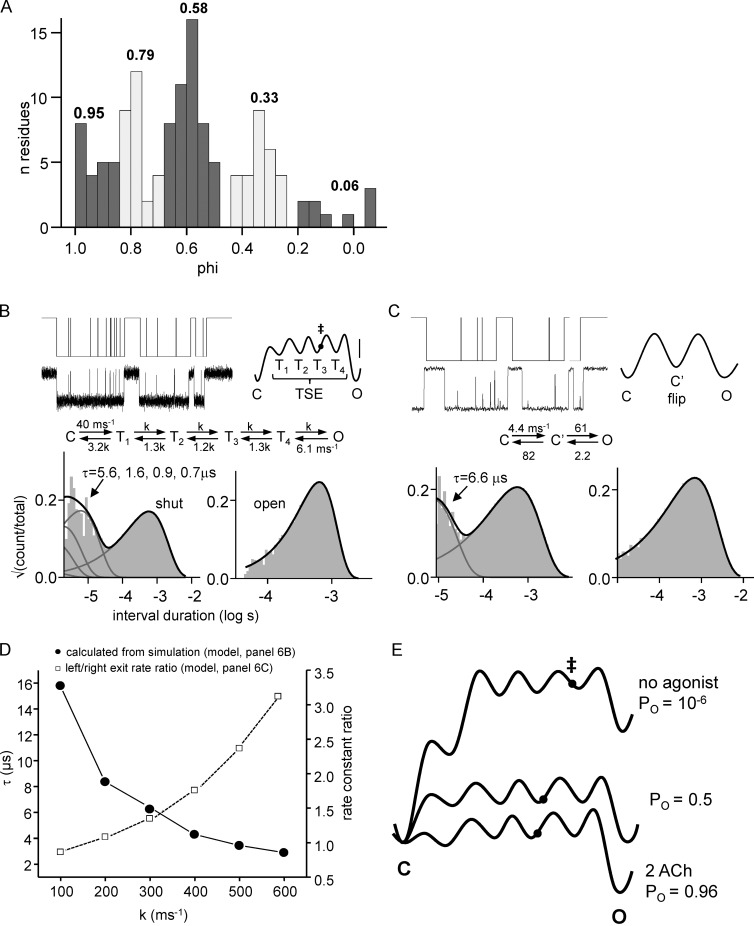
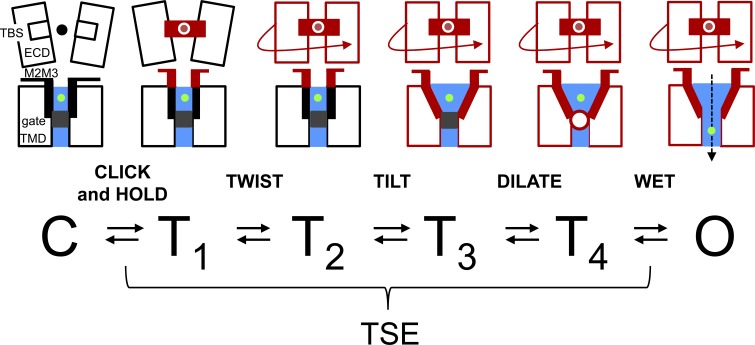
Nicotinic acetylcholine receptors are allosteric proteins that generate membrane currents by isomerizing ("gating") between resting and active conformations under the influence of neurotransmitters. Here, to explore the mechanisms that link the transmitter-binding sites (TBSs) with the distant gate, we use mutant cycle analyses to measure coupling between residue pairs, phi value analyses to sequence domain rearrangements, and current simulations to reproduce a microsecond shut component ("flip") apparent in single-channel recordings. Significant interactions between amino acids separated by >15 Å are rare; an exception is between the αM2-M3 linkers and the TBSs that are ∼30 Å apart. Linker residues also make significant, local interactions within and between subunits. Phi value analyses indicate that without agonists, the linker is the first region in the protein to reach the gating transition state. Together, the phi pattern and flip component suggest that a complete, resting↔active allosteric transition involves passage through four brief intermediate states, with brief shut events arising from sojourns in all or a subset. We derive energy landscapes for gating with and without agonists, and propose a structure-based model in which resting→active starts with spontaneous rearrangements of the M2-M3 linkers and TBSs. These conformational changes stabilize a twisted extracellular domain to promote transmembrane helix tilting, gate dilation, and the formation of a "bubble" that collapses to initiate ion conduction. The energy landscapes suggest that twisting is the most energetically unfavorable step in the resting→active conformational change and that the rate-limiting step in the reverse process is bubble formation.
© 2017 Gupta et al.
Figures

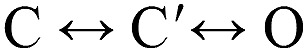
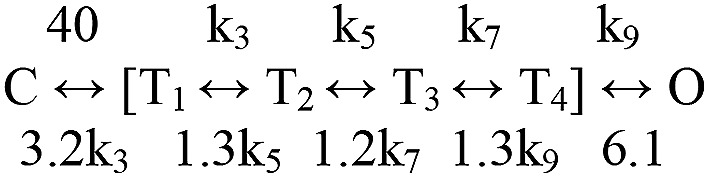
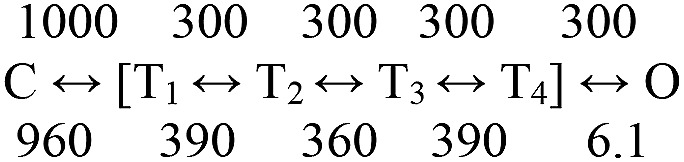
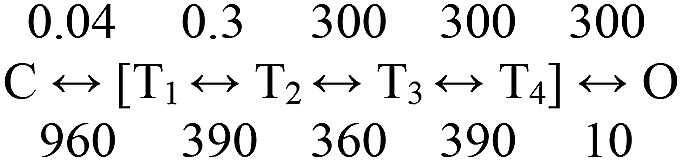
References
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
