

Functional and phenotypic heterogeneity of group 3 innate lymphoid cells
- PMID: 27935637
- PMCID: PMC5290240
- DOI: 10.1111/imm.12697
Functional and phenotypic heterogeneity of group 3 innate lymphoid cells
Abstract
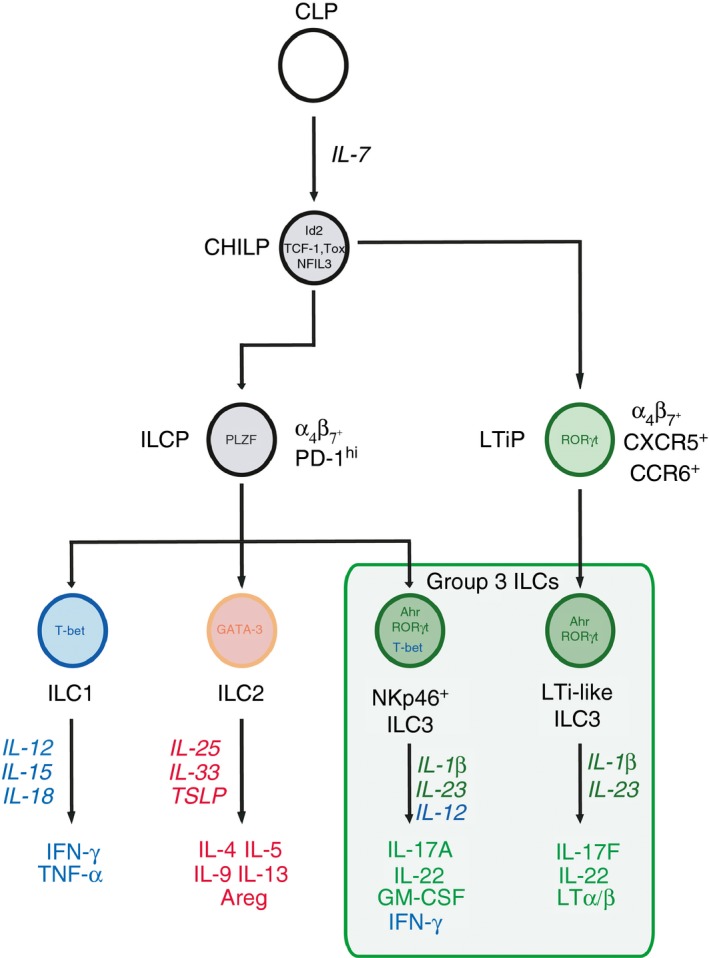
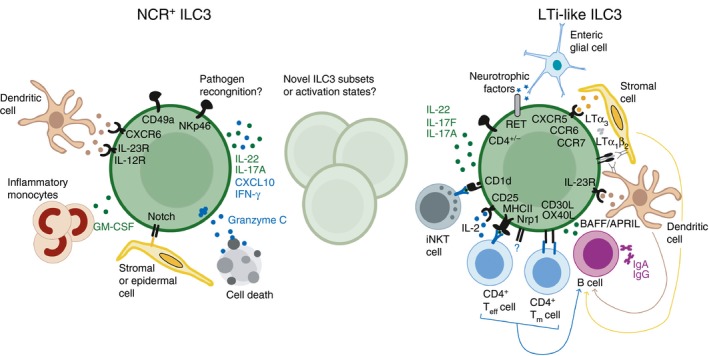
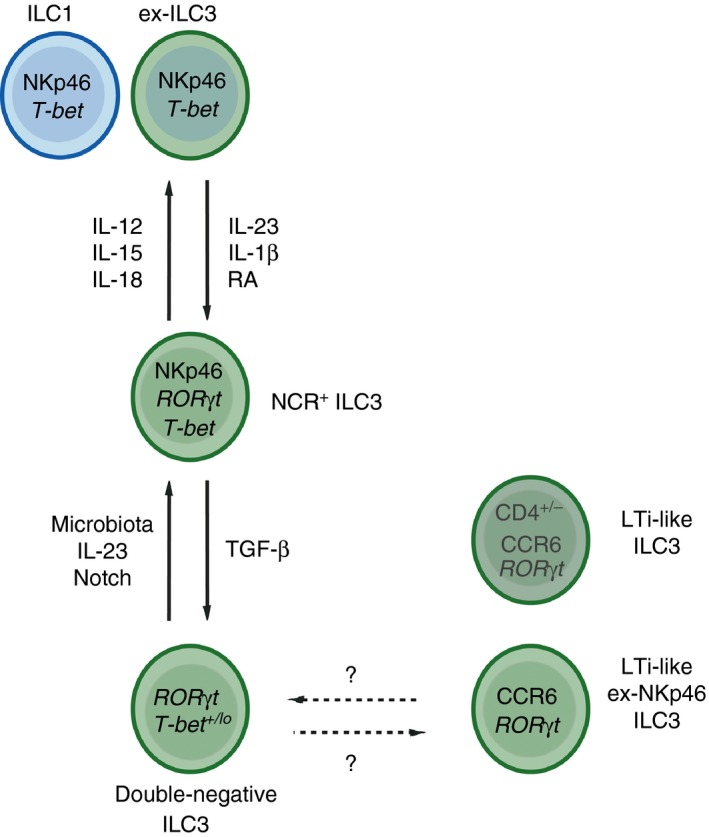
Group 3 innate lymphoid cells (ILC3), defined by expression of the transcription factor retinoid-related orphan receptor γt, play key roles in the regulation of inflammation and immunity in the gastrointestinal tract and associated lymphoid tissues. ILC3 consist largely of two major subsets, NCR+ ILC3 and LTi-like ILC3, but also demonstrate significant plasticity and heterogeneity. Recent advances have begun to dissect the relationship between ILC3 subsets and to define distinct functional states within the intestinal tissue microenvironment. In this review we discuss the ever-expanding roles of ILC3 in the context of intestinal homeostasis, infection and inflammation - with a focus on comparing and contrasting the relative contributions of ILC3 subsets.
Keywords: group 3 innate lymphoid cells; innate lymphoid cell; lymphoid tissue-inducer cells; mucosal immunology.
© 2016 The Authors. Immunology published by John Wiley & Sons Ltd.
Figures
References
-
- Klose CS, Artis D. Innate lymphoid cells as regulators of immunity, inflammation and tissue homeostasis. Nat Immunol 2016; 17:765–74. - PubMed
-
- Walker JA, Barlow JL, McKenzie AN. Innate lymphoid cells – how did we miss them? Nat Rev Immunol 2013; 13:75–87. - PubMed
-
- Spits H, Artis D, Colonna M, Diefenbach A, Di Santo JP, Eberl G et al Innate lymphoid cells – a proposal for uniform nomenclature. Nat Rev Immunol 2013; 13:145–9. - PubMed
-
- Klose CS, Flach M, Mohle L, Rogell L, Hoyler T, Ebert K et al Differentiation of type 1 ILCs from a common progenitor to all helper‐like innate lymphoid cell lineages. Cell 2014; 157:340–56. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
