

Gene Expression and Silencing Studies in Phytophthora infestans Reveal Infection-Specific Nutrient Transporters and a Role for the Nitrate Reductase Pathway in Plant Pathogenesis
- PMID: 27936244
- PMCID: PMC5176271
- DOI: 10.1371/journal.ppat.1006097
Gene Expression and Silencing Studies in Phytophthora infestans Reveal Infection-Specific Nutrient Transporters and a Role for the Nitrate Reductase Pathway in Plant Pathogenesis
Abstract
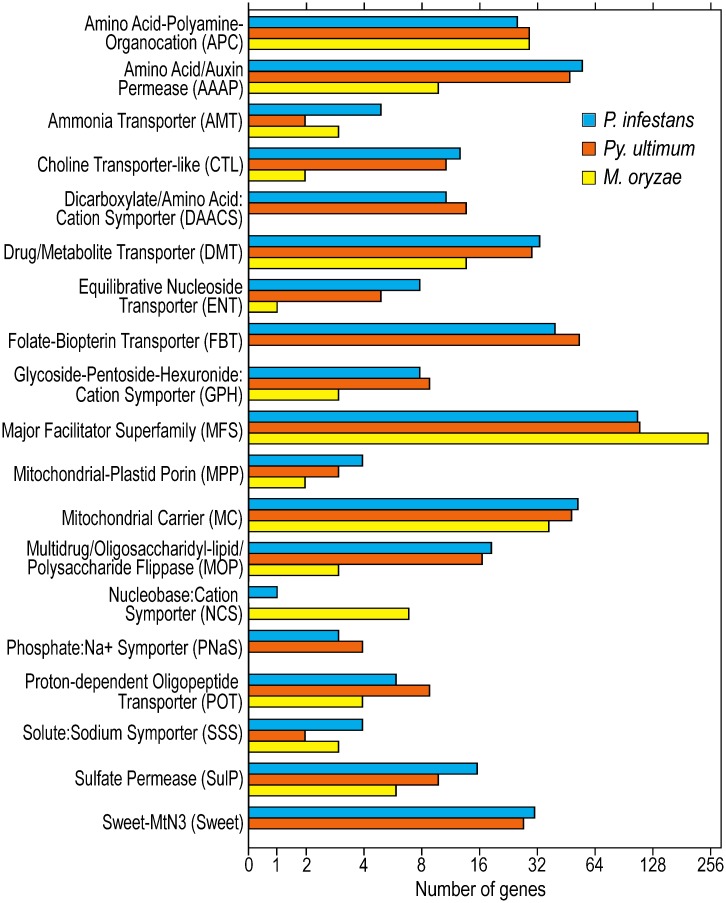
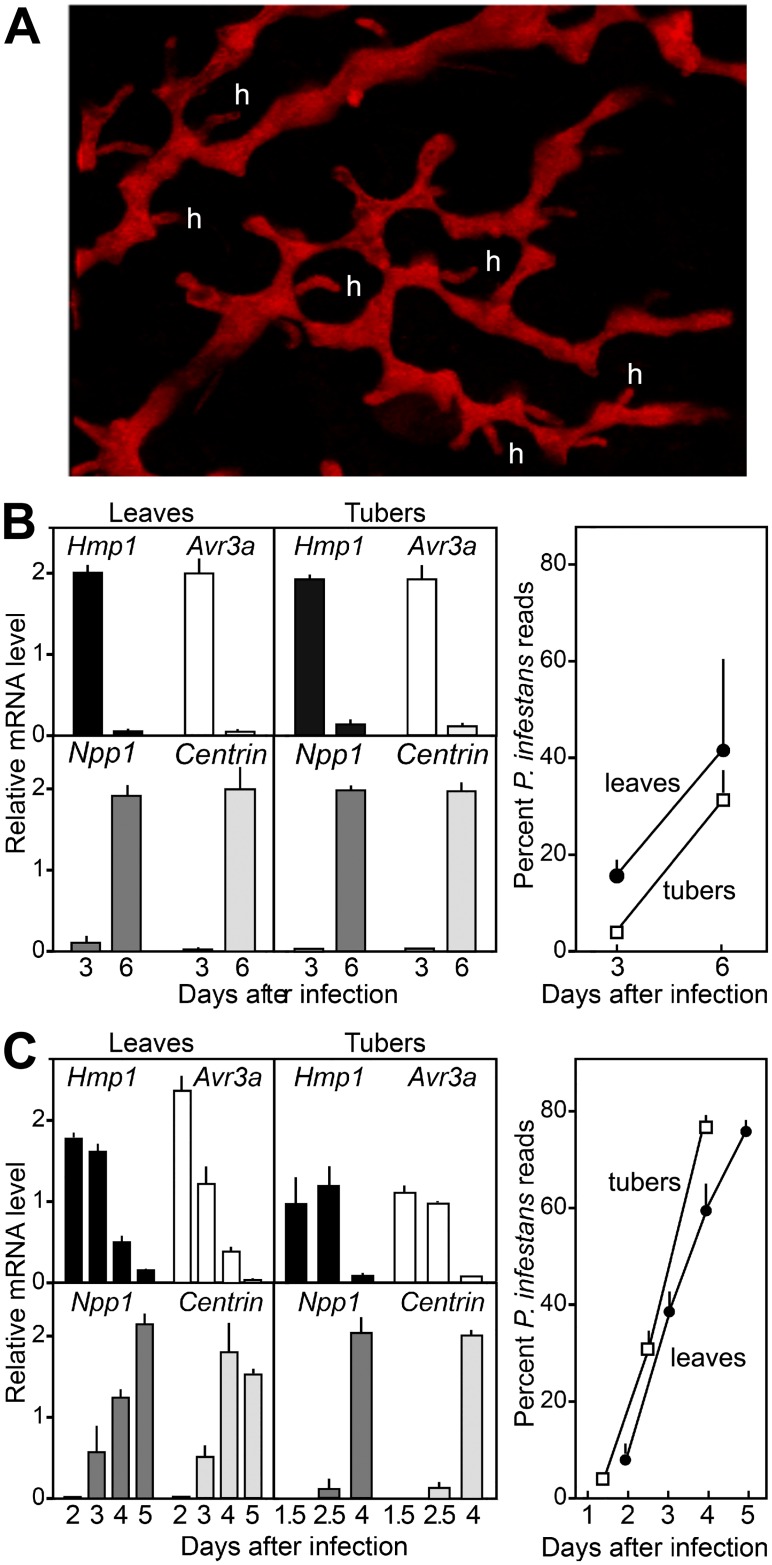
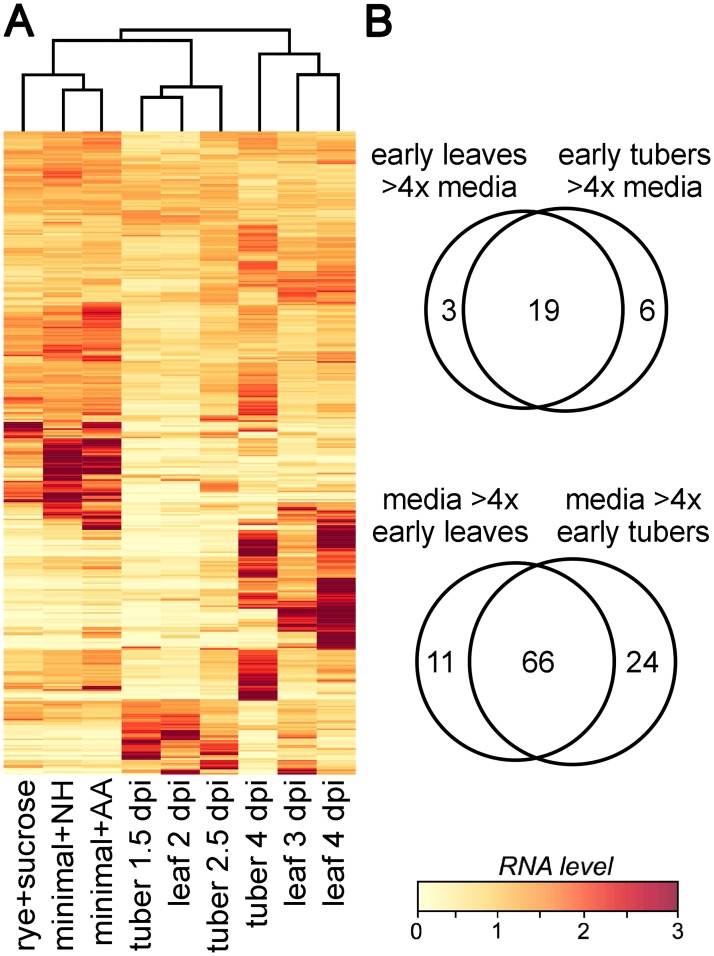
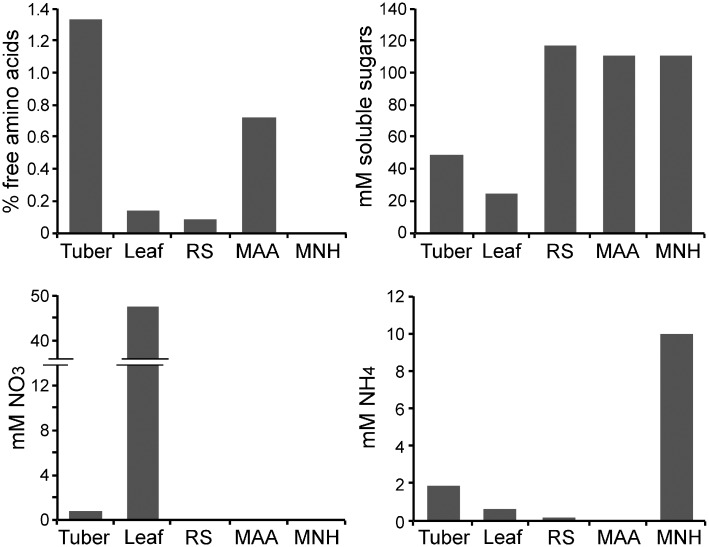
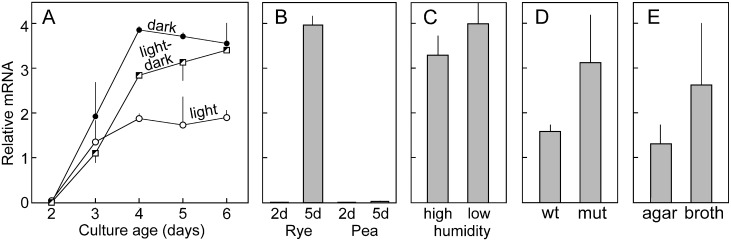
To help learn how phytopathogens feed from their hosts, genes for nutrient transporters from the hemibiotrophic potato and tomato pest Phytophthora infestans were annotated. This identified 453 genes from 19 families. Comparisons with a necrotrophic oomycete, Pythium ultimum var. ultimum, and a hemibiotrophic fungus, Magnaporthe oryzae, revealed diversity in the size of some families although a similar fraction of genes encoded transporters. RNA-seq of infected potato tubers, tomato leaves, and several artificial media revealed that 56 and 207 transporters from P. infestans were significantly up- or down-regulated, respectively, during early infection timepoints of leaves or tubers versus media. About 17 were up-regulated >4-fold in both leaves and tubers compared to media and expressed primarily in the biotrophic stage. The transcription pattern of many genes was host-organ specific. For example, the mRNA level of a nitrate transporter (NRT) was about 100-fold higher during mid-infection in leaves, which are nitrate-rich, than in tubers and three types of artificial media, which are nitrate-poor. The NRT gene is physically linked with genes encoding nitrate reductase (NR) and nitrite reductase (NiR), which mobilize nitrate into ammonium and amino acids. All three genes were coregulated. For example, the three genes were expressed primarily at mid-stage infection timepoints in both potato and tomato leaves, but showed little expression in potato tubers. Transformants down-regulated for all three genes were generated by DNA-directed RNAi, with silencing spreading from the NR target to the flanking NRT and NiR genes. The silenced strains were nonpathogenic on leaves but colonized tubers. We propose that the nitrate assimilation genes play roles both in obtaining nitrogen for amino acid biosynthesis and protecting P. infestans from natural or fertilization-induced nitrate and nitrite toxicity.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

References
-
- Yi M, Valent B. Communication between filamentous pathogens and plants at the biotrophic interface. Ann Rev Phytopathol. 2013;51:587–611. - PubMed
-
- Fernandez J, Marroquin-Guzman M, Wilson RA. Mechanisms of nutrient acquisition and utilization during fungal infections of leaves. Ann Rev Phytopathol. 2014;52:155–174. - PubMed
-
- Hahn M, Neef U, Struck C, Gottfert M, Mendgen K. A putative amino acid transporter is specifically expressed in haustoria of the rust fungus Uromyces fabae. Molec Plant Microbe Interact. 1997;10:438–445. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
