

Neuronal migration during development and the amyloid precursor protein
- PMID: 27939704
- PMCID: PMC5157842
- DOI: 10.1016/j.cois.2016.08.001
Neuronal migration during development and the amyloid precursor protein
Abstract
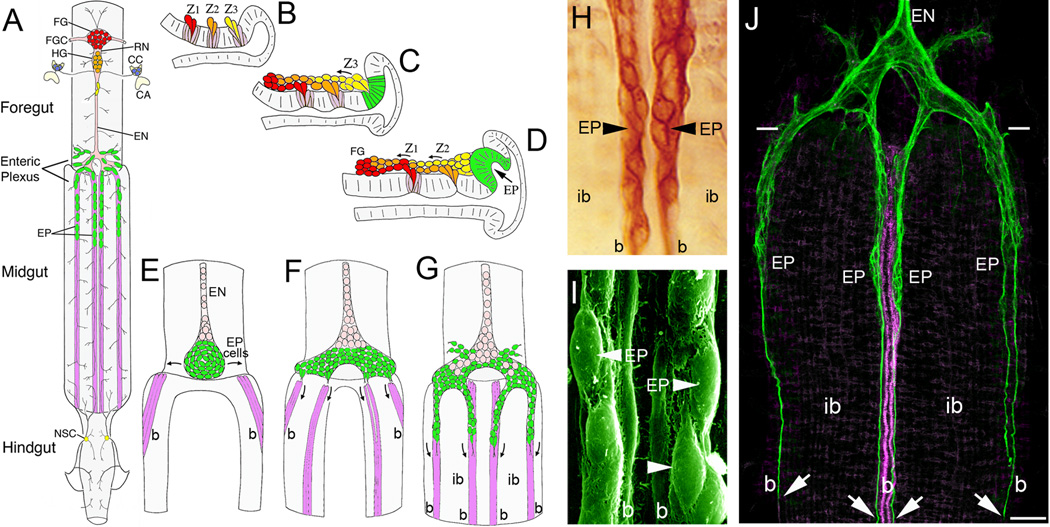
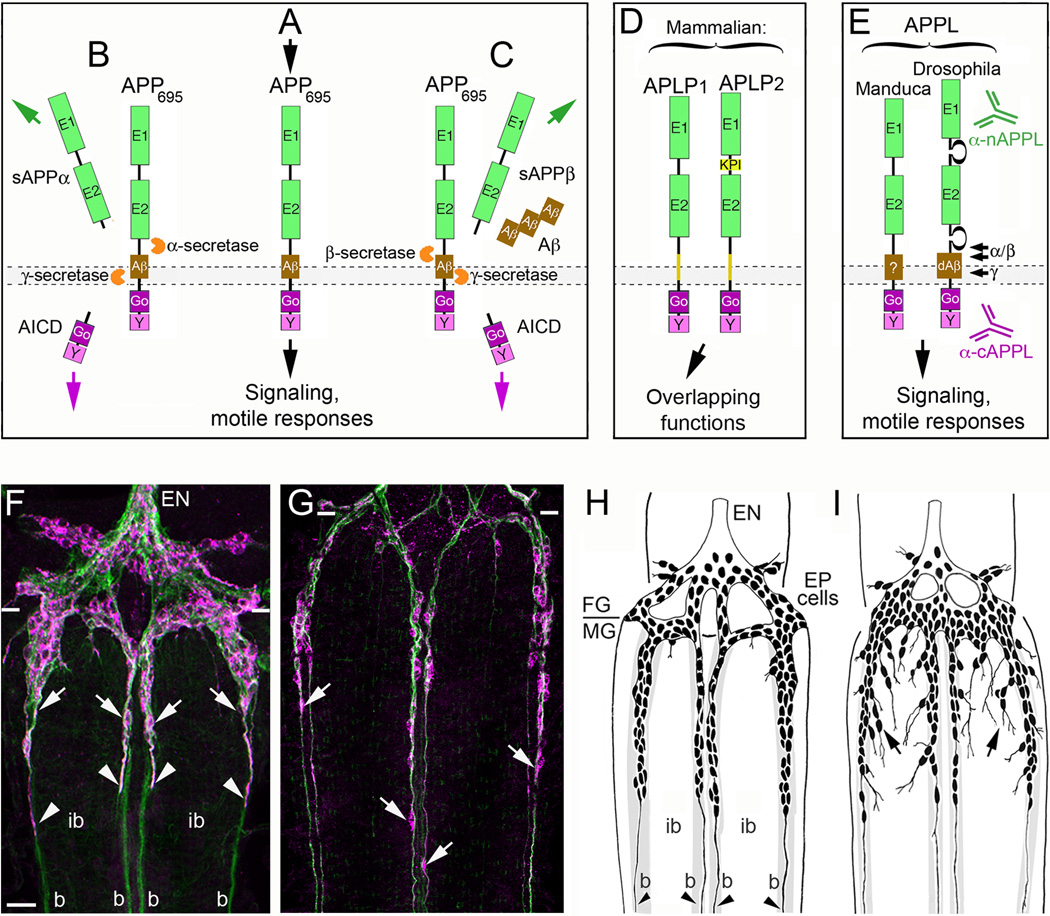
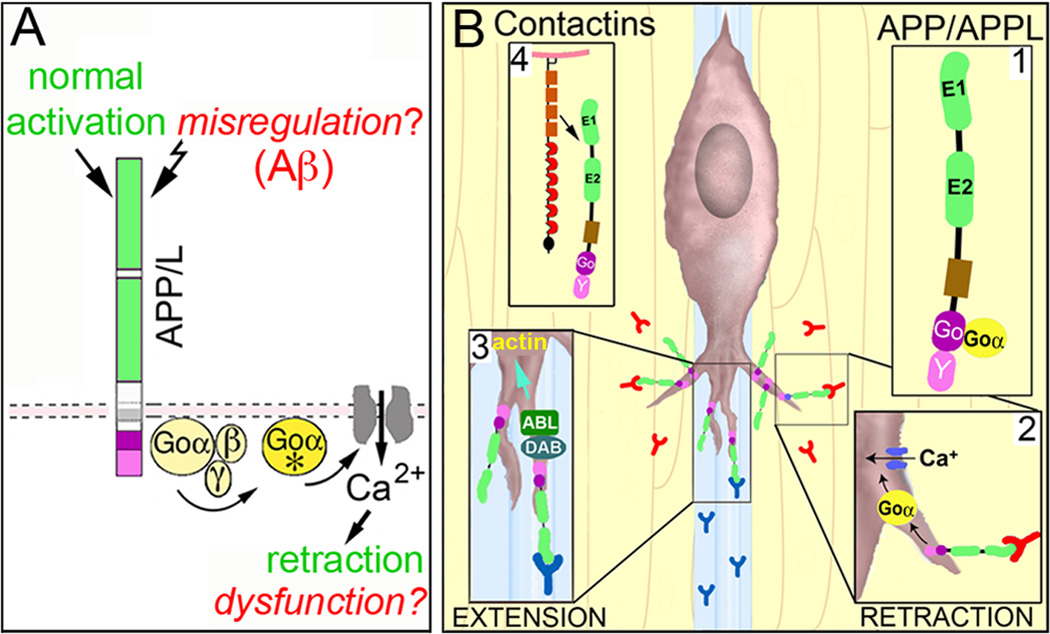
The Amyloid Precursor Protein (APP) is the source of amyloid peptides that accumulate in Alzheimer's disease. However, members of the APP family are strongly expressed in the developing nervous systems of invertebrates and vertebrates, where they regulate neuronal guidance, synaptic remodeling, and injury responses. In contrast to mammals, insects express only one APP ortholog (APPL), simplifying investigations into its normal functions. Recent studies have shown that APPL regulates neuronal migration in the developing insect nervous system, analogous to the roles ascribed to APP family proteins in the mammalian cortex. The comparative simplicity of insect systems offers new opportunities for deciphering the signaling mechanisms by which this enigmatic class of proteins contributes to the formation and function of the nervous system.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures
References
-
- Feng Y, Walsh CA. Protein-protein interactions, cytoskeletal regulation and neuronal migration. Nat Rev Neurosci. 2001;2:408–416. - PubMed
-
- Hatten ME. New directions in neuronal migration. Science. 2002;297:1660–1663. - PubMed
-
- Cajal Ramón y . Studies in Vertebrate Neurogenesis. Springfield, Ill: Thomas, Charles C; 1929.
-
- Le Douarin NM, Kalcheim C. The Neural Crest. Cambridge, England: Cambridge U. Press; 1999.
-
-
Gertz CC, Kriegstein AR. Neuronal Migration Dynamics in the Developing Ferret Cortex. J Neurosci. 2015;35:14307–14315. This study used advanced neuronal labeling and imaging methods to investigate how migratory patterns are different in gyrencephalic (folded) brains versus lissencephalic (smooth) brains, providing new insight into the types of migratory errors that might contribute to congenital defects in human brain development.
-
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
