

Unique aspects of the developing lung circulation: structural development and regulation of vasomotor tone
- PMID: 27942377
- PMCID: PMC5130074
- DOI: 10.1086/688890
Unique aspects of the developing lung circulation: structural development and regulation of vasomotor tone
Abstract
This review summarizes our current knowledge on lung vasculogenesis and angiogenesis during normal lung development and the regulation of fetal and postnatal pulmonary vascular tone. In comparison to that of the adult, the pulmonary circulation of the fetus and newborn displays many unique characteristics. Moreover, altered development of pulmonary vasculature plays a more prominent role in compromised pulmonary vasoreactivity than in the adult. Clinically, a better understanding of the developmental changes in pulmonary vasculature and vasomotor tone and the mechanisms that are disrupted in disease states can lead to the development of new therapies for lung diseases characterized by impaired alveolar structure and pulmonary hypertension.
Keywords: endothelial; fetal; newborn; pulmonary circulation; smooth muscle.
Figures
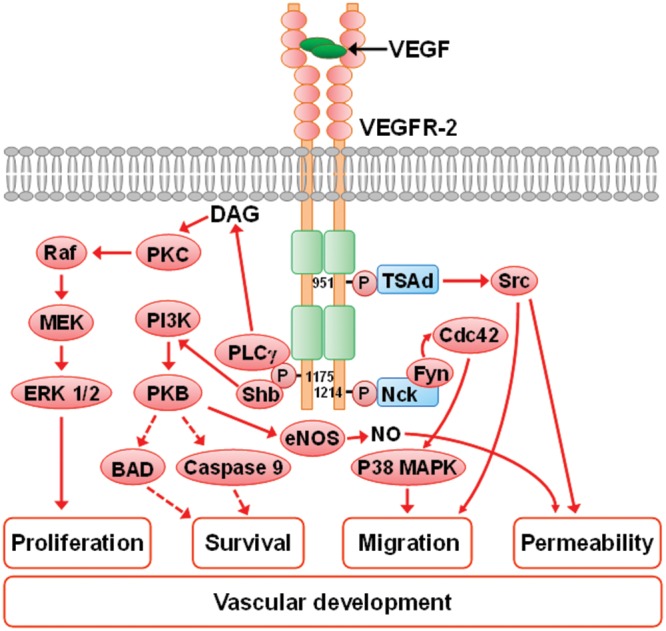
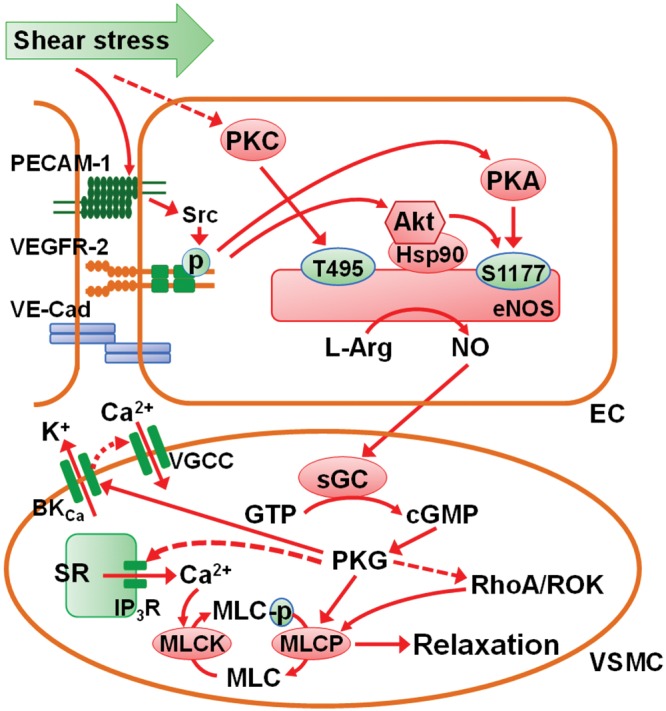
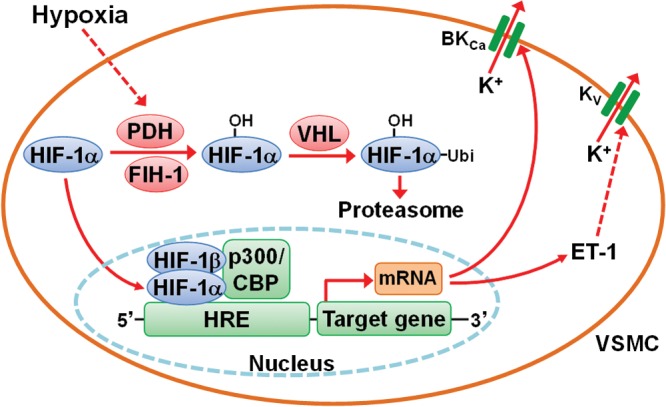
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
