

Donor exosomes rather than passenger leukocytes initiate alloreactive T cell responses after transplantation
- PMID: 27942611
- PMCID: PMC5142759
- DOI: 10.1126/sciimmunol.aaf8759
Donor exosomes rather than passenger leukocytes initiate alloreactive T cell responses after transplantation
Abstract
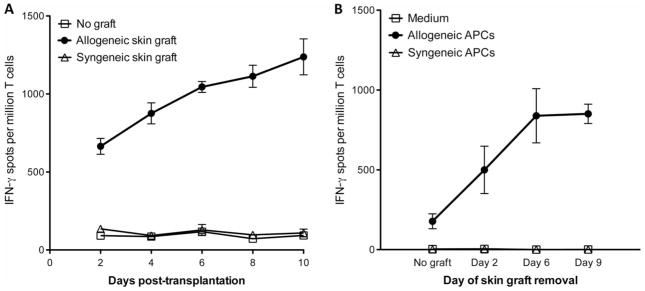
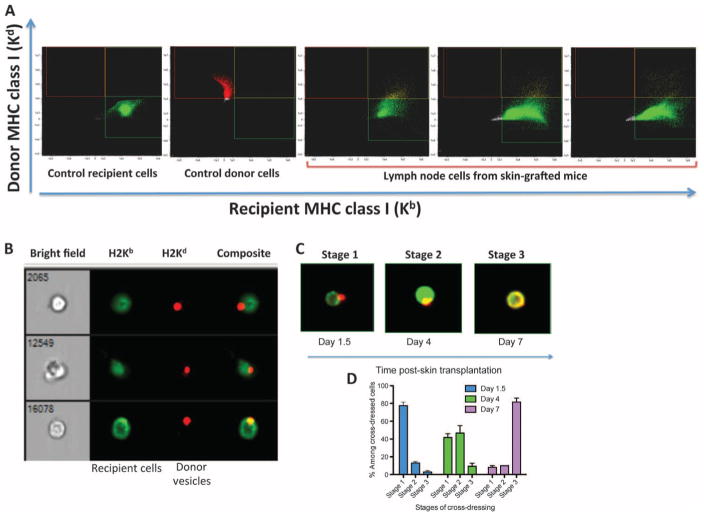
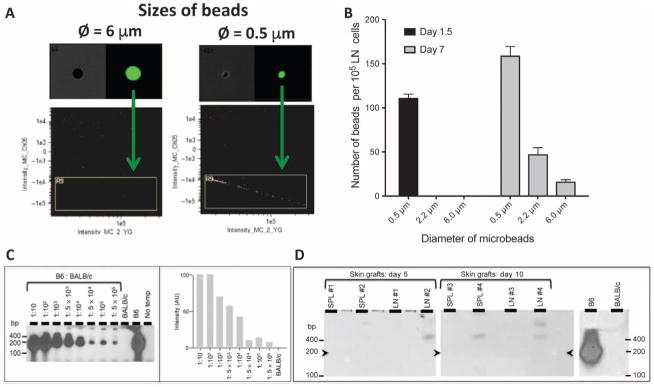
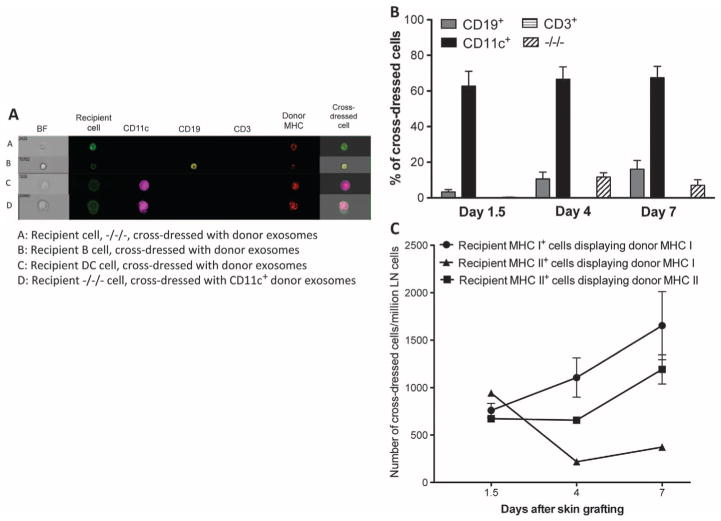
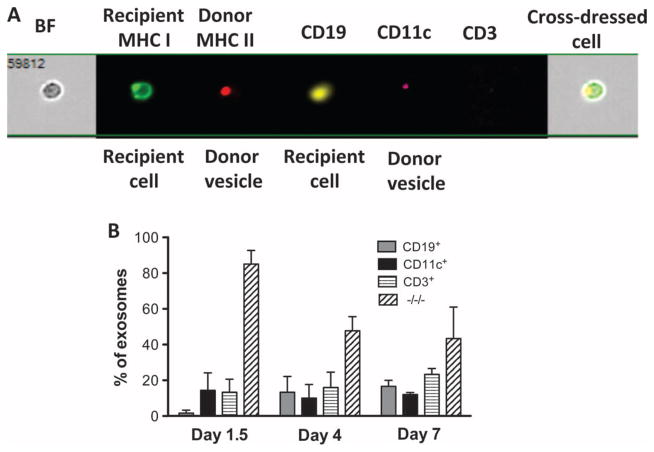
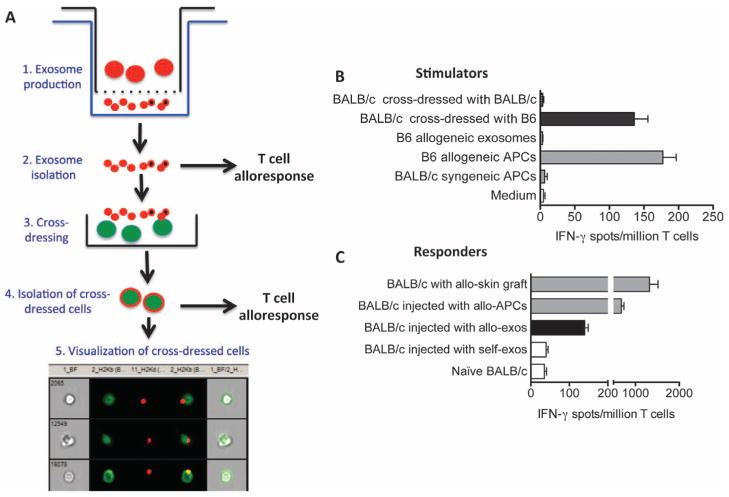
Transplantation of allogeneic organs and tissues represents a lifesaving procedure for a variety of patients affected with end-stage diseases. Although current immunosuppressive therapy prevents early acute rejection, it is associated with nephrotoxicity and increased risks for infection and neoplasia. This stresses the need for selective immune-based therapies relying on manipulation of lymphocyte recognition of donor antigens. The passenger leukocyte theory states that allograft rejection is initiated by recipient T cells recognizing donor major histocompatibility complex (MHC) molecules displayed on graft leukocytes migrating to the host's lymphoid organs. We revisited this concept in mice transplanted with allogeneic skin, heart, or islet grafts using imaging flow cytometry. We observed no donor cells in the lymph nodes and spleen of skin-grafted mice, but we found high numbers of recipient cells displaying allogeneic MHC molecules (cross-dressed) acquired from donor microvesicles (exosomes). After heart or islet transplantation, we observed few donor leukocytes (100 per million) but large numbers of recipient cells cross-dressed with donor MHC (>90,000 per million). Last, we showed that purified allogeneic exosomes induced proinflammatory alloimmune responses by T cells in vitro and in vivo. Collectively, these results suggest that recipient antigen-presenting cells cross-dressed with donor MHC rather than passenger leukocytes trigger T cell responses after allotransplantation.
Conflict of interest statement
The authors declare that they have no competing financial interests.
Figures
Similar articles
-
Donor-derived exosomes: the trick behind the semidirect pathway of allorecognition.Curr Opin Organ Transplant. 2017 Feb;22(1):46-54. doi: 10.1097/MOT.0000000000000372. Curr Opin Organ Transplant. 2017. PMID: 27898464 Free PMC article. Review.
-
Limited T cell response to donor MHC peptides during allograft rejection. Implications for selective immune therapy in transplantation.J Immunol. 1994 Aug 1;153(3):938-45. J Immunol. 1994. PMID: 7517977
-
Presentation of donor major histocompatibility complex antigens by bone marrow dendritic cell-derived exosomes modulates allograft rejection.Transplantation. 2003 Nov 27;76(10):1503-10. doi: 10.1097/01.TP.0000092494.75313.38. Transplantation. 2003. PMID: 14657694
-
Cross-dressing by donor dendritic cells after allogeneic bone marrow transplantation contributes to formation of the immunological synapse and maximizes responses to indirectly presented antigen.J Immunol. 2014 Jun 1;192(11):5426-33. doi: 10.4049/jimmunol.1302490. Epub 2014 Apr 30. J Immunol. 2014. PMID: 24790149
-
Emerging role of exosomes in allorecognition and allograft rejection.Curr Opin Organ Transplant. 2018 Feb;23(1):22-27. doi: 10.1097/MOT.0000000000000489. Curr Opin Organ Transplant. 2018. PMID: 29189413 Free PMC article. Review.
Cited by
-
Dendritic Cells: Versatile Players in Renal Transplantation.Front Immunol. 2021 May 19;12:654540. doi: 10.3389/fimmu.2021.654540. eCollection 2021. Front Immunol. 2021. PMID: 34093544 Free PMC article. Review.
-
T cell antigenicity and immunogenicity of allogeneic exosomes.Am J Transplant. 2021 Jul;21(7):2583-2589. doi: 10.1111/ajt.16591. Epub 2021 Apr 30. Am J Transplant. 2021. PMID: 33794063 Free PMC article.
-
Graft-derived extracellular vesicles transported across subcapsular sinus macrophages elicit B cell alloimmunity after transplantation.Sci Transl Med. 2021 Mar 17;13(585):eabb0122. doi: 10.1126/scitranslmed.abb0122. Sci Transl Med. 2021. PMID: 33731430 Free PMC article.
-
Extracellular Vesicles in chondrogenesis and Cartilage regeneration.J Cell Mol Med. 2021 Jun;25(11):4883-4892. doi: 10.1111/jcmm.16290. Epub 2021 May 4. J Cell Mol Med. 2021. PMID: 33942981 Free PMC article. Review.
-
Engineering dendritic cell vaccines to improve cancer immunotherapy.Nat Commun. 2019 Nov 27;10(1):5408. doi: 10.1038/s41467-019-13368-y. Nat Commun. 2019. PMID: 31776331 Free PMC article. Review.
References
-
- Lechler R, Lombardi G. Structural aspects of allorecognition. Curr Opin Immunol. 1991;3:715–721. - PubMed
-
- Suchin EJ, Langmuir PB, Palmer E, Sayegh MH, Wells AD, Turka LA. Quantifying the frequency of alloreactive T cells in vivo: New answers to an old question. J Immunol. 2001;166:973–981. - PubMed
-
- Benichou G, Fedoseyeva E, Lehmann PV, Olson CA, Geysen HM, McMillan M, Sercarz EE. Limited T cell response to donor MHC peptides during allograft rejection. Implications for selective immune therapy in transplantation. J Immunol. 1994;153:938–945. - PubMed
-
- Liu Z, Sun YK, Xi YP, Hong B, Harris PE, Reed EF, Suciu-Foca N. Limited usage of T cell receptor V beta genes by allopeptide-specific T cells. J Immunol. 1993;150:3180–3186. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
