

Epigenetic Regulation of the Sex Determination Gene MeGI in Polyploid Persimmon
- PMID: 27956470
- PMCID: PMC5240738
- DOI: 10.1105/tpc.16.00532
Epigenetic Regulation of the Sex Determination Gene MeGI in Polyploid Persimmon
Abstract
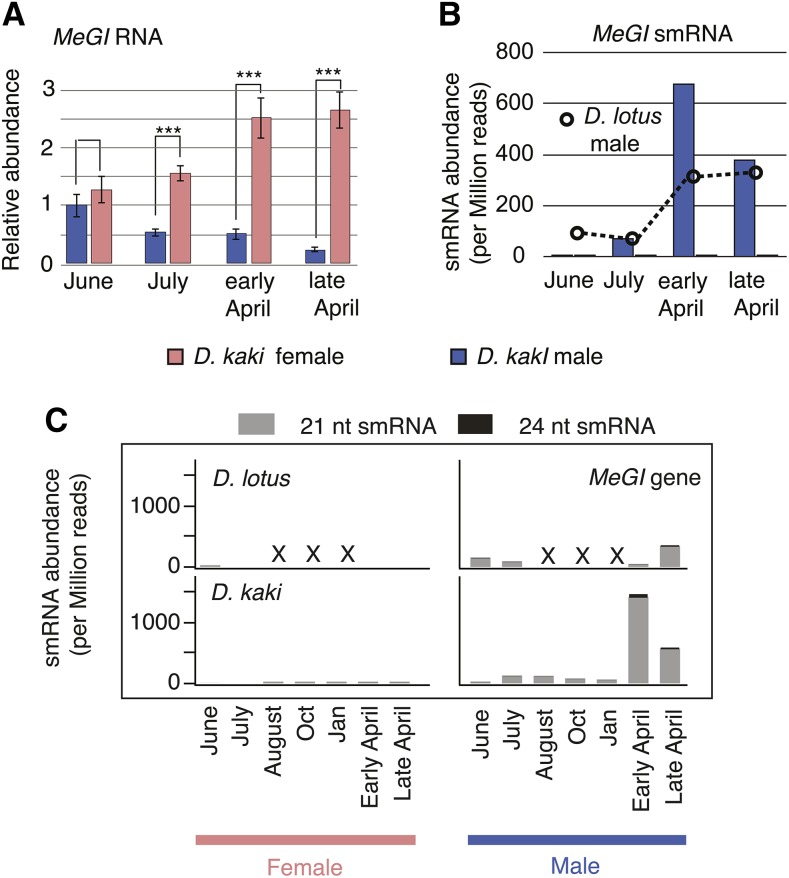
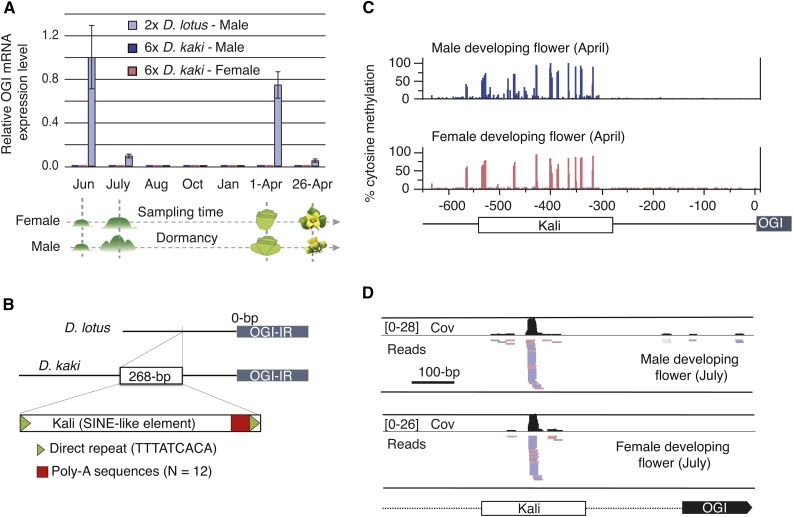
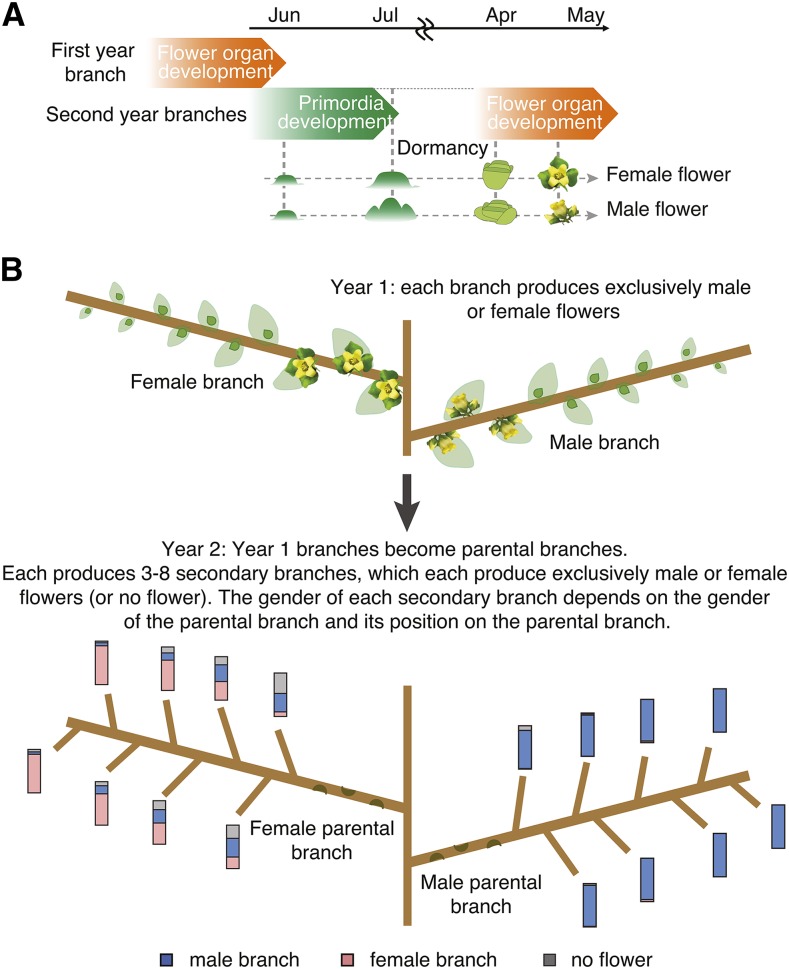
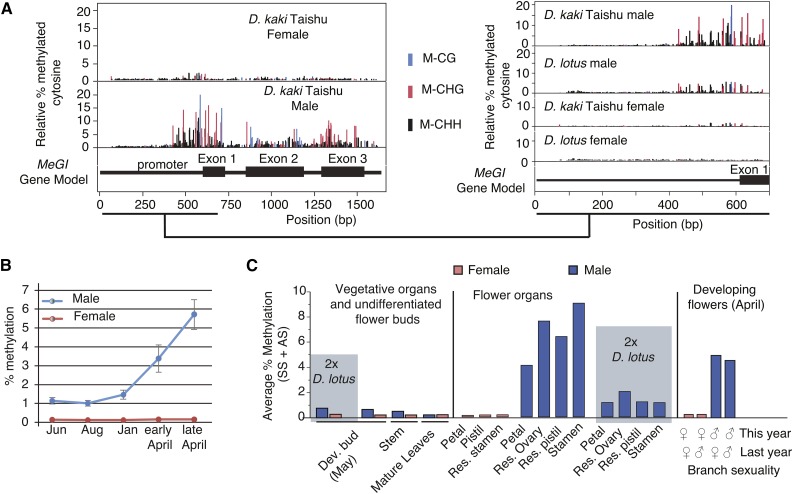
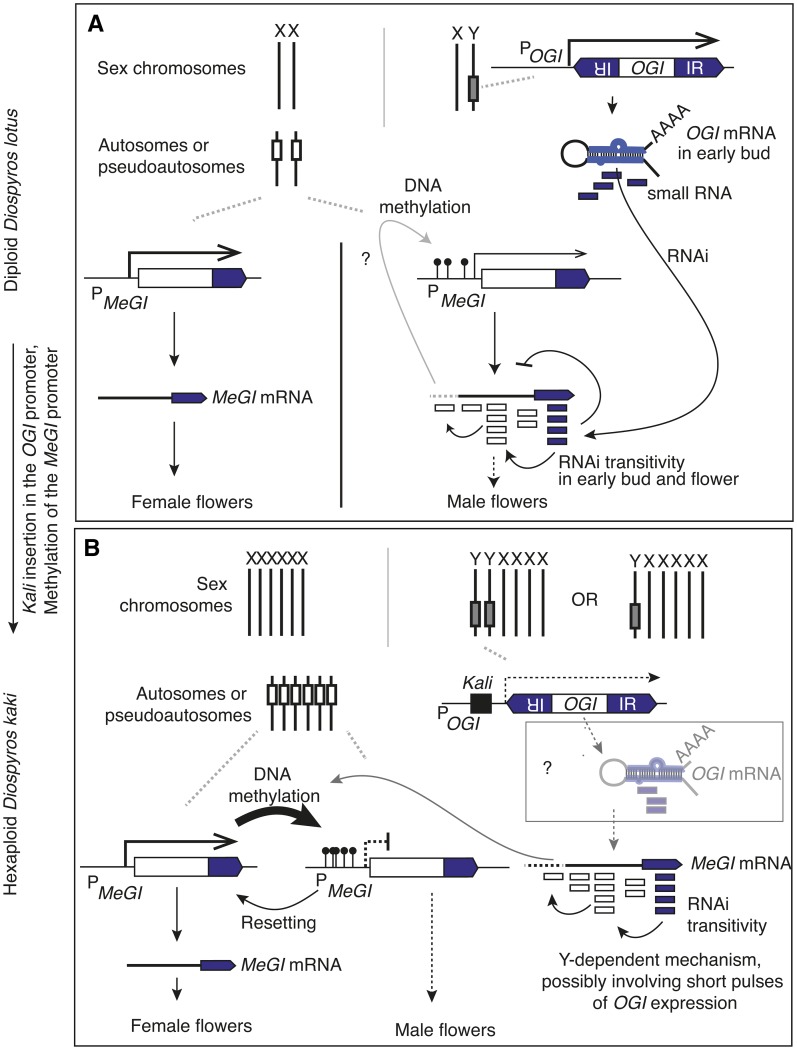
Epigenetic regulation can add a flexible layer to genetic variation, potentially enabling long-term but reversible cis-regulatory changes to an allele while maintaining its DNA sequence. Here, we present a case in which alternative epigenetic states lead to reversible sex determination in the hexaploid persimmon Diospyros kaki Previously, we elucidated the molecular mechanism of sex determination in diploid persimmon and demonstrated the action of a Y-encoded sex determinant pseudogene called OGI, which produces small RNAs targeting the autosomal gene MeGI, resulting in separate male and female individuals (dioecy). We contrast these findings with the discovery, in hexaploid persimmon, of an additional layer of regulation in the form of DNA methylation of the MeGI promoter associated with the production of both male and female flowers in genetically male trees. Consistent with this model, developing male buds exhibited higher methylation levels across the MeGI promoter than developing female flowers from either monoecious or female trees. Additionally, a DNA methylation inhibitor induced developing male buds to form feminized flowers. Concurrently, in Y-chromosome-carrying trees, the expression of OGI is silenced by the presence of a SINE (short interspersed nuclear element)-like insertion in the OGI promoter. Our findings provide an example of an adaptive scenario involving epigenetic plasticity.
© 2016 American Society of Plant Biologists. All rights reserved.
Figures

Comment in
-
The Power of Plasticity in Polyploid Persimmon.Plant Cell. 2016 Dec;28(12):2885-2886. doi: 10.1105/tpc.16.00936. Epub 2016 Dec 15. Plant Cell. 2016. PMID: 27980004 Free PMC article. No abstract available.
References
-
- Akagi T., Henry I.M., Tao R., Comai L. (2014a). Plant genetics. A Y-chromosome-encoded small RNA acts as a sex determinant in persimmons. Science 346: 646–650. - PubMed
-
- Akagi T., Kibe T., Morimura H., Kajita K., Tsujimoto T., Nishiyama S., Kawai T., Yamane H., Tao R. (2014b). Development of molecular markers associated with sexuality in Diospyros lotus L. and their application in D. kaki Thunb. J. Jpn. Soc. Hortic. Sci. 76: 214–221.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
