

Role of CCL7 in Type I Hypersensitivity Reactions in Murine Experimental Allergic Conjunctivitis
- PMID: 27956527
- PMCID: PMC5220120
- DOI: 10.4049/jimmunol.1502416
Role of CCL7 in Type I Hypersensitivity Reactions in Murine Experimental Allergic Conjunctivitis
Abstract
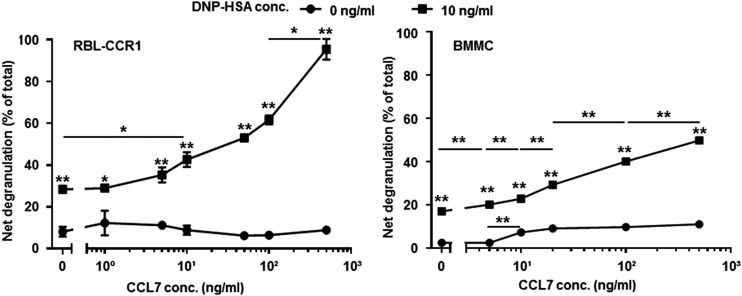
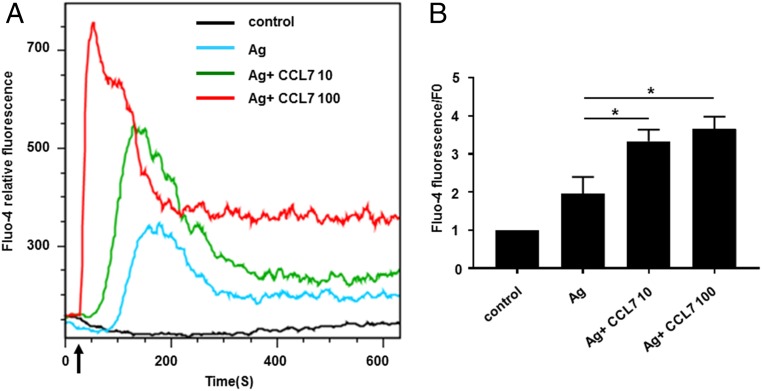
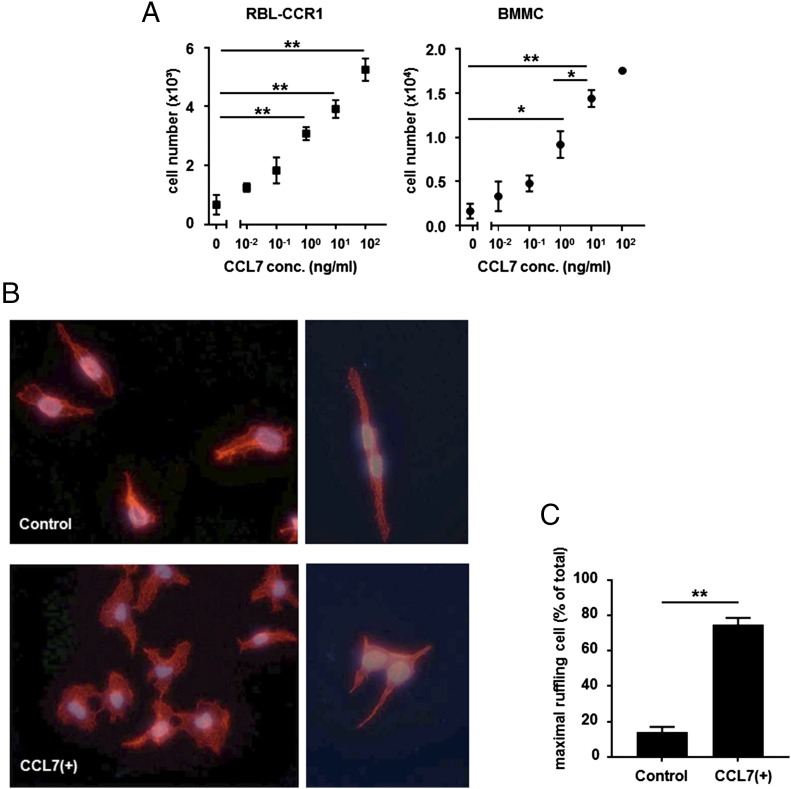
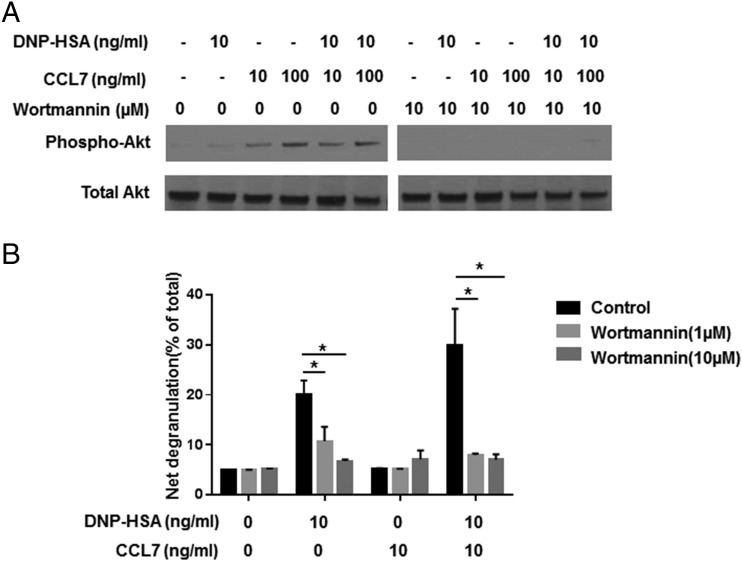
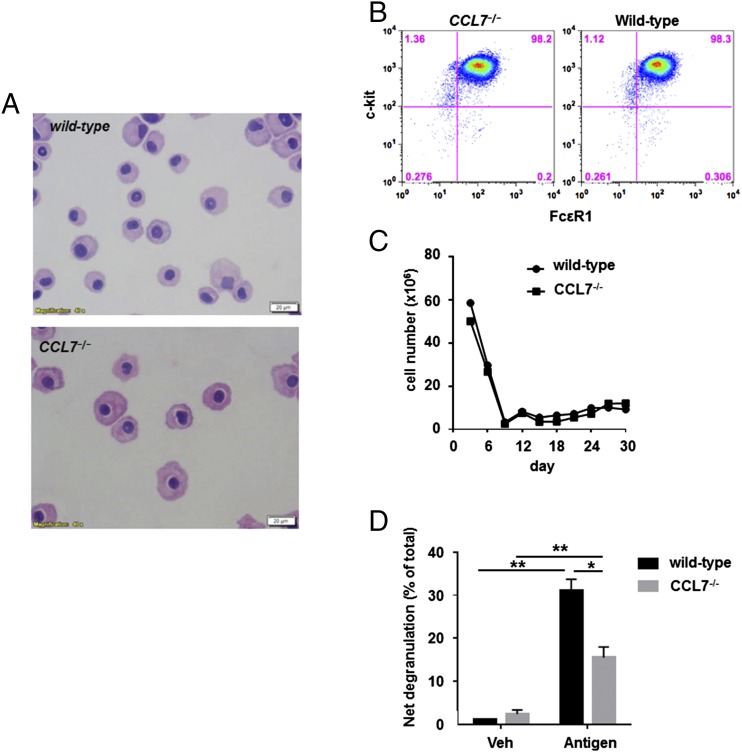
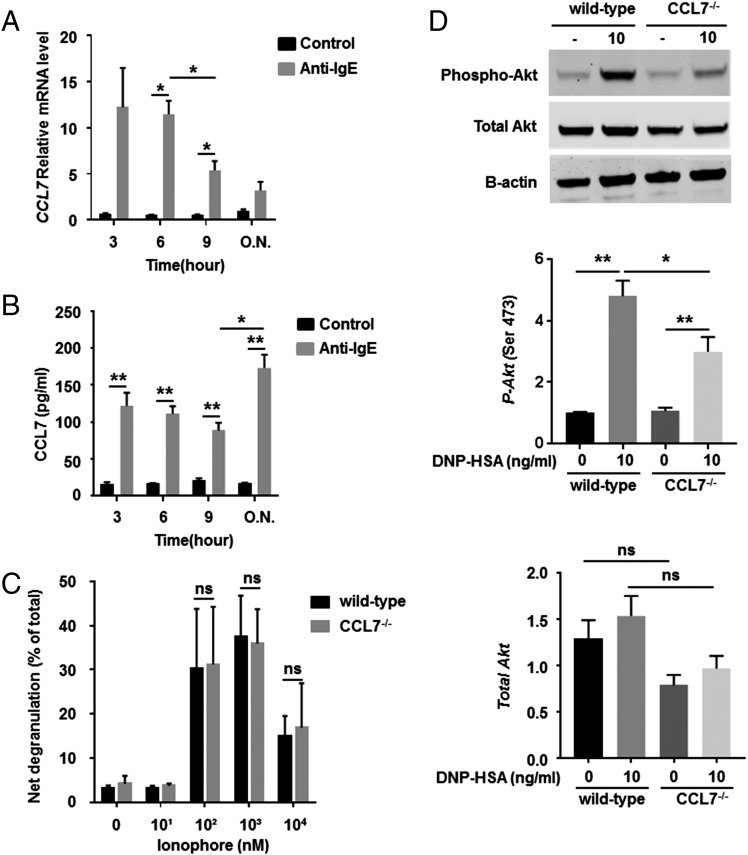
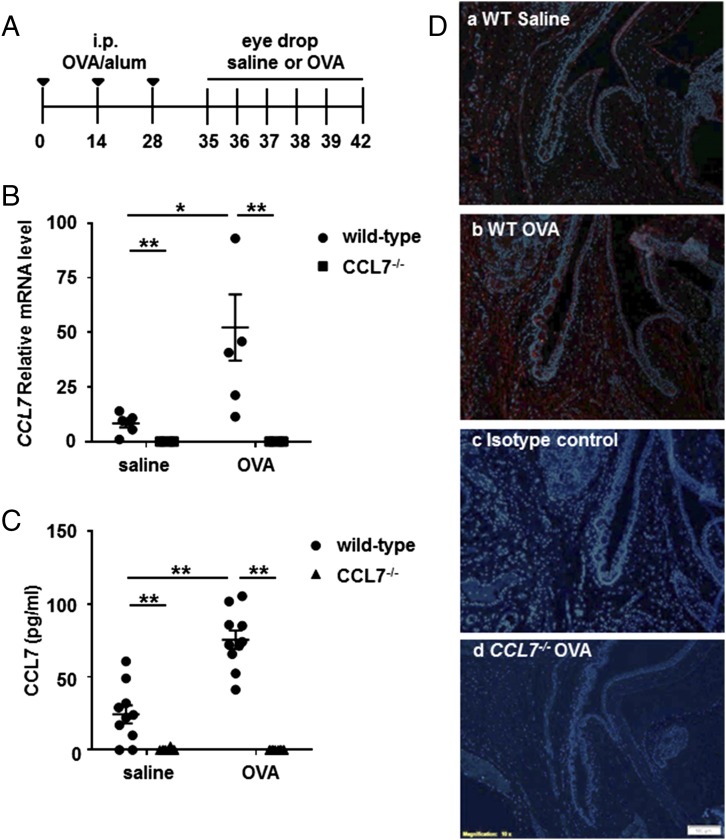
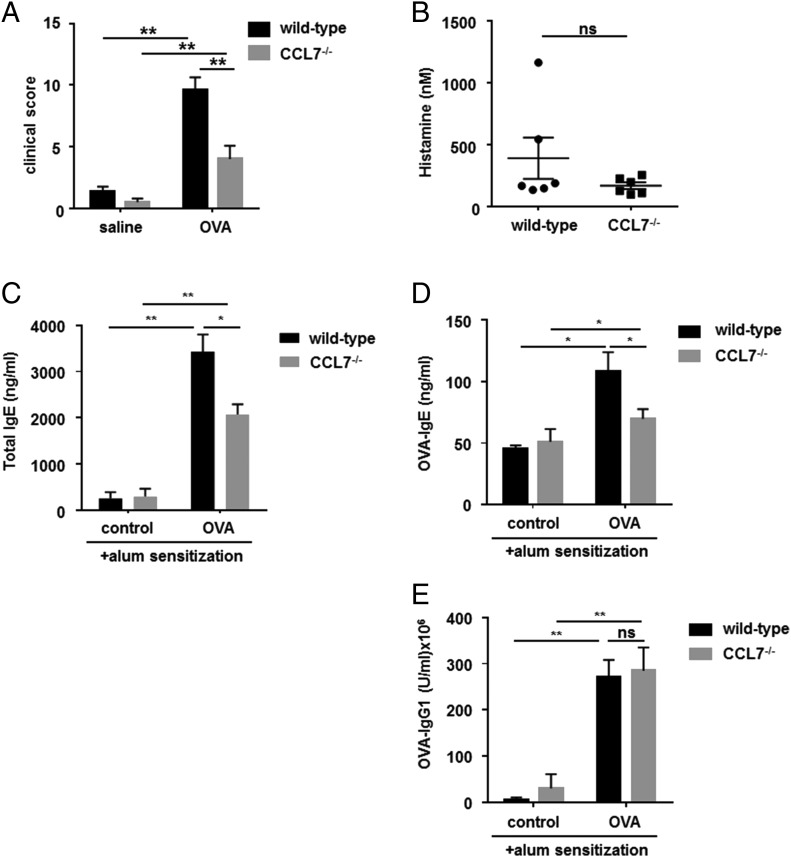
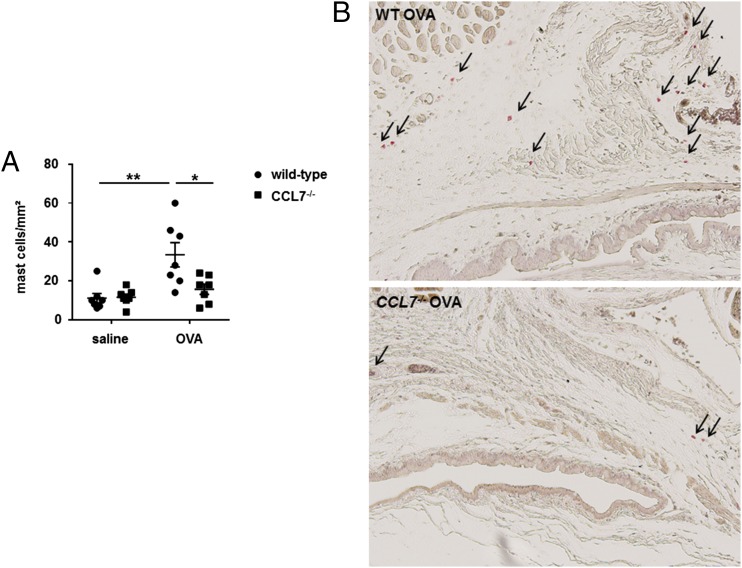
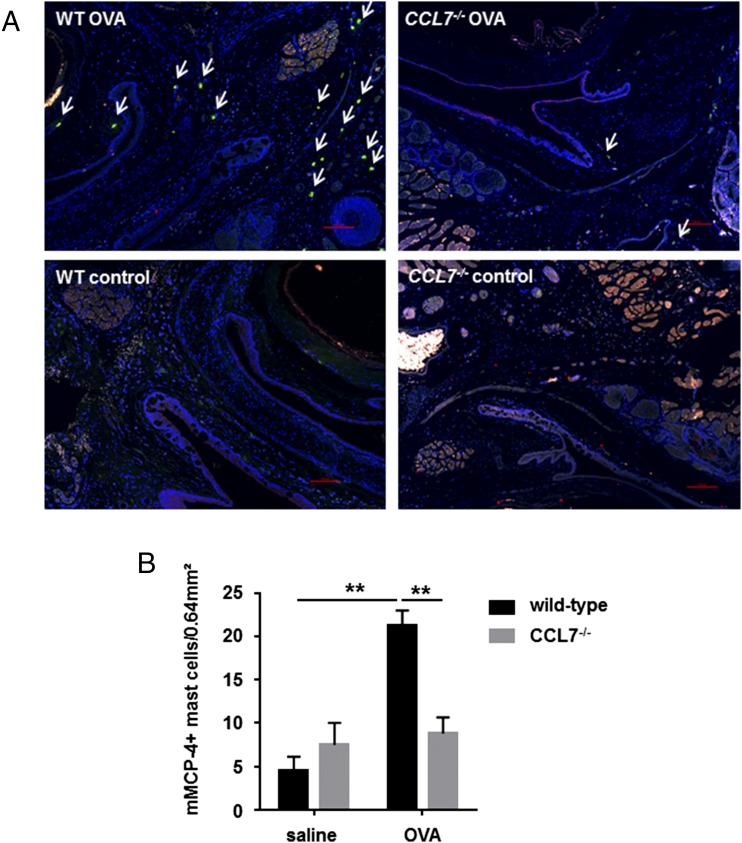
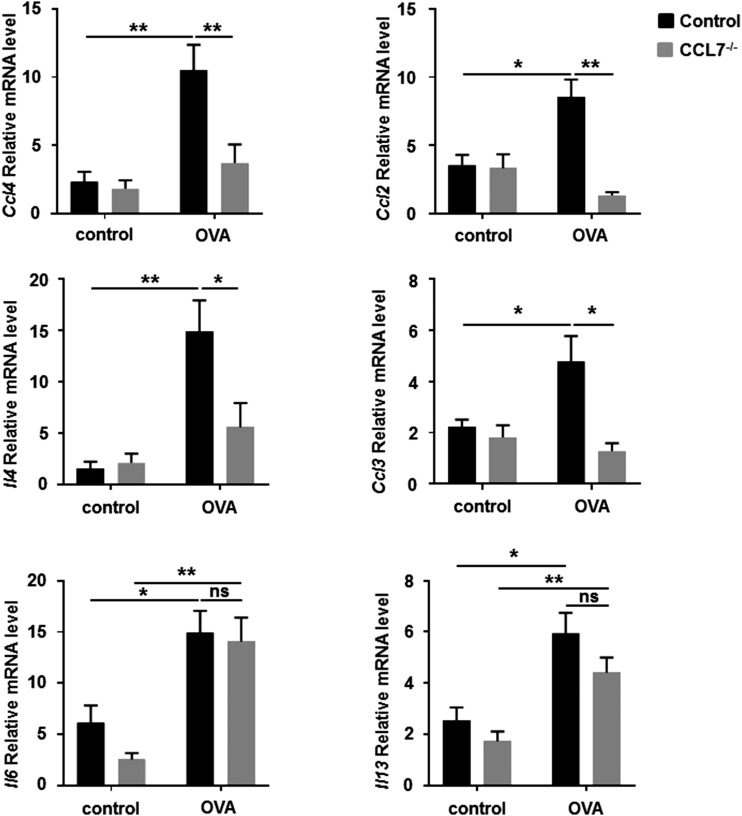
Molecules that are necessary for ocular hypersensitivity reactions include the receptors CCR1 and CCR3; CCL7 is a ligand for these receptors. Therefore, we explored the role of CCL7 in mast cell activity and motility in vitro and investigated the requirement for CCL7 in a murine model of IgE-mediated allergic conjunctivitis. For mast cells treated with IgE and Ag, the presence of CCL7 synergistically enhanced degranulation and calcium influx. CCL7 also induced chemotaxis in mast cells. CCL7-deficient bone marrow-derived mast cells showed decreased degranulation following IgE and Ag treatment compared with wild-type bone marrow-derived mast cells, but there was no difference in degranulation when cells were activated via an IgE-independent pathway. In vivo, CCL7 was upregulated in conjunctival tissue during an OVA-induced allergic response. Notably, the early-phase clinical symptoms in the conjunctiva after OVA challenge were significantly higher in OVA-sensitized wild-type mice than in control challenged wild-type mice; the increase was suppressed in CCL7-deficient mice. In the OVA-induced allergic response, the numbers of conjunctival mast cells were lower in CCL7-deficient mice than in wild-type mice. Our results demonstrate that CCL7 is required for maximal OVA-induced ocular anaphylaxis, mast cell recruitment in vivo, and maximal FcεRI-mediated mast cell activation in vitro. A better understanding of the role of CCL7 in mediating ocular hypersensitivity reactions will provide insights into mast cell function and novel treatments for allergic ocular diseases.
Copyright © 2017 by The American Association of Immunologists, Inc.
Figures
References
-
- O’Brien T. P. 2013. Allergic conjunctivitis: an update on diagnosis and management. Curr. Opin. Allergy Clin. Immunol. 13: 543–549. - PubMed
-
- Gilfillan A. M., Tkaczyk C. 2006. Integrated signalling pathways for mast-cell activation. Nat. Rev. Immunol. 6: 218–230. - PubMed
-
- Toda M., Nakamura T., Ohbayashi M., Ikeda Y., Dawson M., Aye C. C., Miyazaki D., Ono S. J. 2007. Mechanisms of leukocyte trafficking in allergic diseases: insights into new therapies targeting chemokines and chemokine receptors. Expert Rev. Clin. Immunol. 3: 351–364. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
