

Efficient Breeding by Genomic Mating
- PMID: 27965707
- PMCID: PMC5126051
- DOI: 10.3389/fgene.2016.00210
Efficient Breeding by Genomic Mating
Abstract
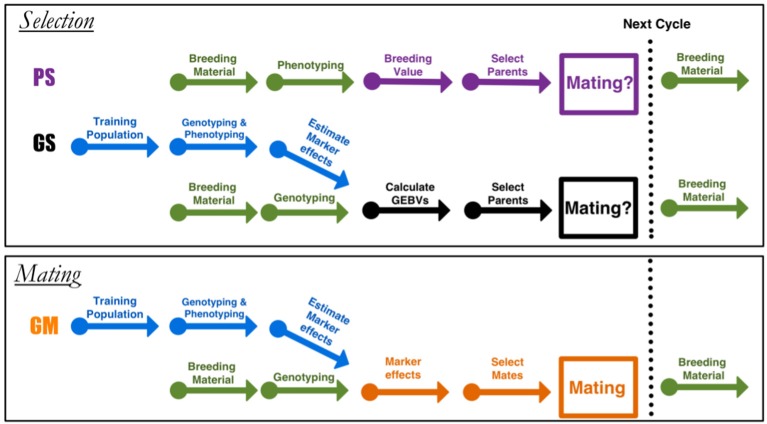
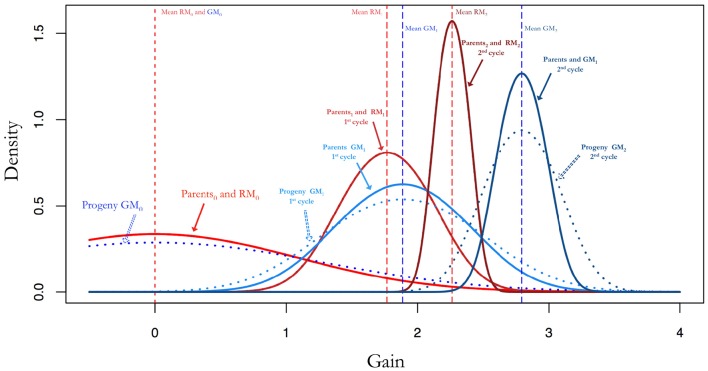
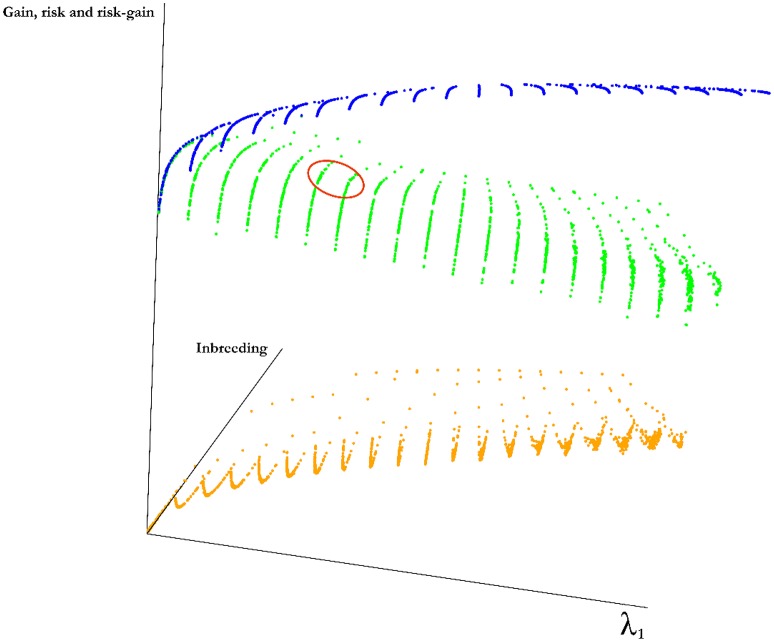
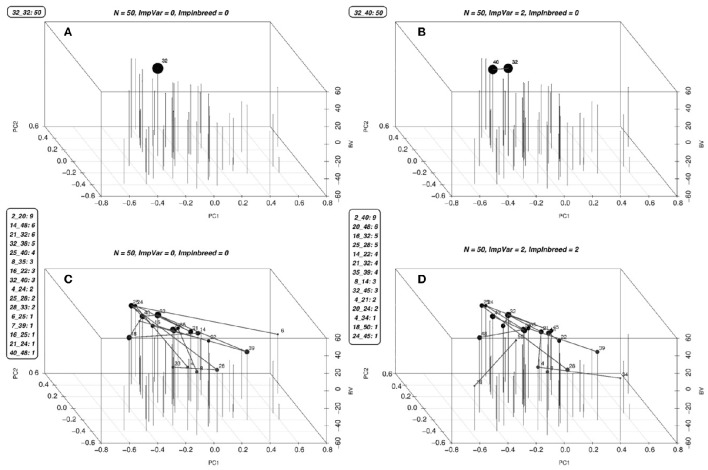
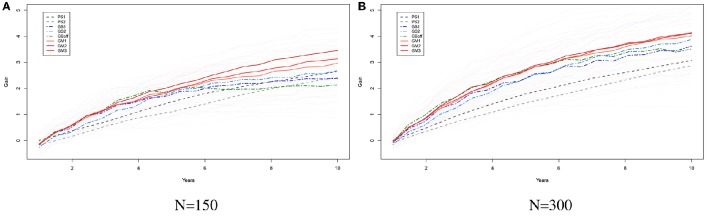
Selection in breeding programs can be done by using phenotypes (phenotypic selection), pedigree relationship (breeding value selection) or molecular markers (marker assisted selection or genomic selection). All these methods are based on truncation selection, focusing on the best performance of parents before mating. In this article we proposed an approach to breeding, named genomic mating, which focuses on mating instead of truncation selection. Genomic mating uses information in a similar fashion to genomic selection but includes information on complementation of parents to be mated. Following the efficiency frontier surface, genomic mating uses concepts of estimated breeding values, risk (usefulness) and coefficient of ancestry to optimize mating between parents. We used a genetic algorithm to find solutions to this optimization problem and the results from our simulations comparing genomic selection, phenotypic selection and the mating approach indicate that current approach for breeding complex traits is more favorable than phenotypic and genomic selection. Genomic mating is similar to genomic selection in terms of estimating marker effects, but in genomic mating the genetic information and the estimated marker effects are used to decide which genotypes should be crossed to obtain the next breeding population.
Keywords: breeding; complex traits; genome-wide markers; genomic selection; phenotypic selection.
Figures
References
-
- Akdemir D., Godfrey O. U. (2015). EMMREML: Fitting Mixed Models with Known Covariance Structures. R package version 3.1.
-
- Berg P., Nielsen J., Sørensen M. K. (2006). Eva: realized and predicted optimal genetic contributions, in Proceedings of the 8th World Congress on Genetics Applied to Livestock Production, Belo Horizonte, Minas Gerais, Brazil, 13-18 August (Porto Alegre, RS: Instituto Prociência; ).
-
- Bernardo R., Charcosset A. (2006). Usefulness of gene information in marker-assisted recurrent selection: a simulation appraisal. Crop Sci. 46, 614–621. 10.2135/cropsci2005.05-0088 - DOI
LinkOut - more resources
Full Text Sources
Other Literature Sources