

A pseudouridine synthase module is essential for mitochondrial protein synthesis and cell viability
- PMID: 27974379
- PMCID: PMC5210091
- DOI: 10.15252/embr.201643391
A pseudouridine synthase module is essential for mitochondrial protein synthesis and cell viability
Abstract
Pseudouridylation is a common post-transcriptional modification in RNA, but its functional consequences at the cellular level remain largely unknown. Using a proximity-biotinylation assay, we identified a protein module in mitochondrial RNA granules, platforms for post-transcriptional RNA modification and ribosome assembly, containing several proteins of unknown function including three uncharacterized pseudouridine synthases, TRUB2, RPUSD3, and RPUSD4. TRUB2 and RPUSD4 were previously identified as core essential genes in CRISPR/Cas9 screens. Depletion of the individual enzymes produced specific mitochondrial protein synthesis and oxidative phosphorylation assembly defects without affecting mitochondrial mRNA levels. Investigation of the molecular targets in mitochondrial RNA by pseudouridine-Seq showed that RPUSD4 plays a role in the pseudouridylation of a single residue in the 16S rRNA, a modification that is essential for its stability and assembly into the mitochondrial ribosome, while TRUB2/RPUSD3 were similarly involved in pseudouridylating specific residues in mitochondrial mRNAs. These results establish essential roles for epitranscriptomic modification of mitochondrial RNA in mitochondrial protein synthesis, oxidative phosphorylation, and cell survival.
Keywords: epitranscriptomic modification; mitochondrial protein synthesis; oxidative phosphorylation; pseudouridine synthase; ribosome assembly.
© 2016 The Authors.
Figures
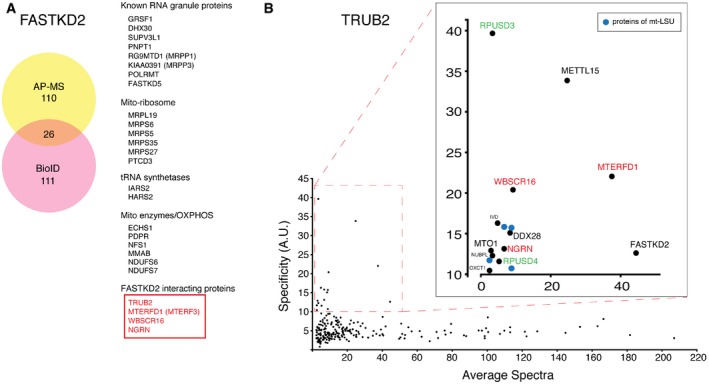
Identification of FASTKD2 interacting proteins by AP–MS and BioID. 26 high‐confidence interactors detected by both techniques are listed on the right.
Prey specificity graph for BioID interactome of TRUB2 protein. Prey specificity was calculated as the relative enrichment of interaction of individual preys and TRUB2, compared to their interaction with 138 other baits (42 mitochondrial baits, 96 baits from other cellular compartments).
- A
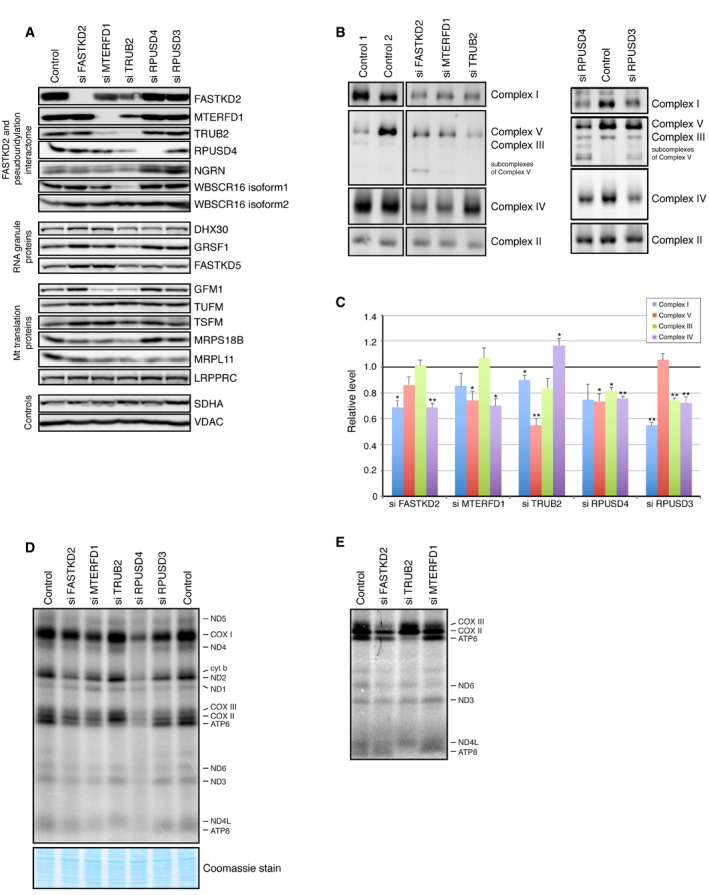
Immunoblot analysis of indicated proteins in control and siRNA‐treated cells.
- B
BN–PAGE analysis of siRNA‐mediated depletion shows an OXPHOS defect as revealed by subunit‐specific antibodies against individual OXPHOS complexes.
- C
Quantification of the levels of individual OXPHOS complexes normalized to complex II levels. The graph represents the relative abundance of individual complexes in cells treated with the specified siRNA versus controls. The bars represent the mean ± SEM of 3–7 independent experiments. P‐values were calculated using paired two‐tailed t‐test (*P < 0.05; **P < 0.01).
- D, E
Pulse‐labeling mitochondrial translation experiment of the 13 mitochondria‐encoded polypeptides (seven subunits of complex I [ND], three subunits of complex IV [COX], two subunits of complex V [ATP], and one subunit of complex III [cyt b]) in control and siRNA‐treated cells. (E) Detail of a mitochondrial pulse‐labeling experiment showing a severe decrease in the translation of ATP6 and ATP8 in TRUB2‐depleted cells.
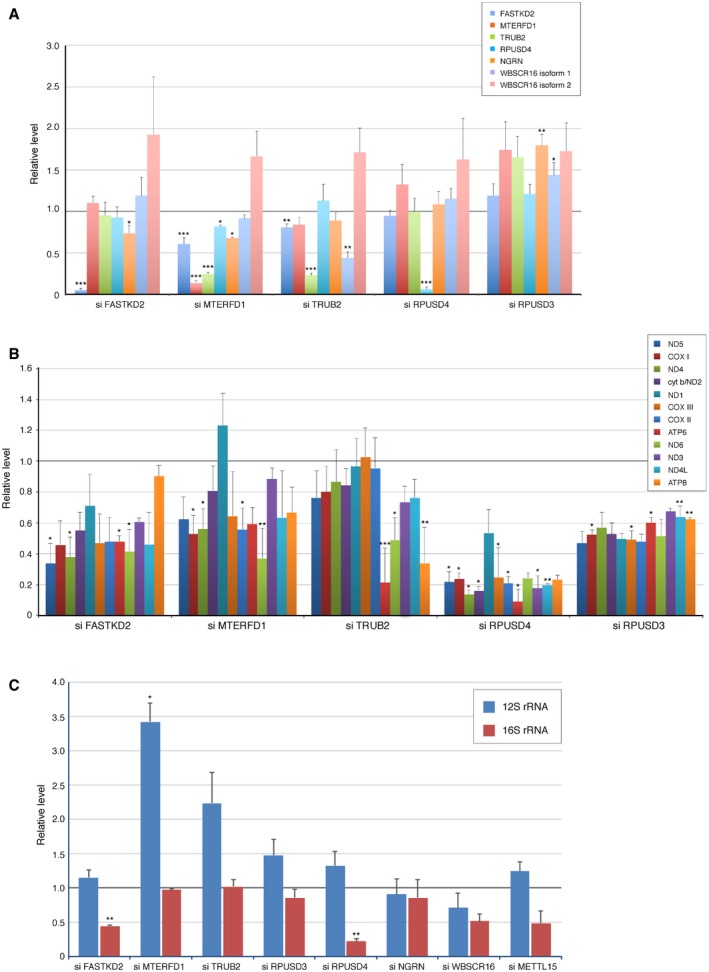
Quantification of the levels of individual pseudouridine synthase interacting proteins normalized to SDHA in cells treated with siRNA. The graph represents the relative protein levels compared to controls. The bars represent mean ± SEM of 4–13 independent experiments.
Quantification of the synthesis of individual mitochondria‐encoded polypeptides in siRNA‐treated cells. The graph represents the relative levels compared to controls. The bars represent mean ± SEM of 2–4 independent experiments.
qRT–PCR analysis of 12S and 16S rRNA in siRNA‐treated cells. The graph represents the relative levels compared to controls. The bars represent mean ± SEM of 2–3 independent experiments.
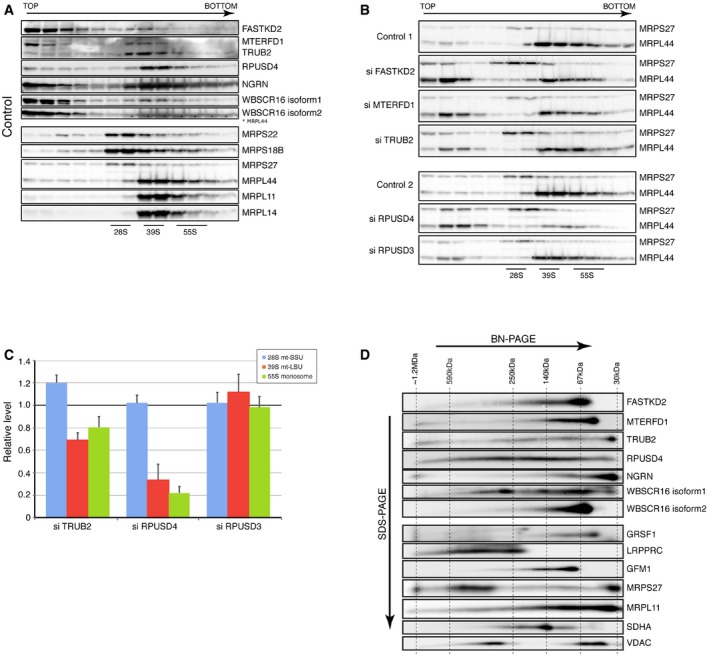
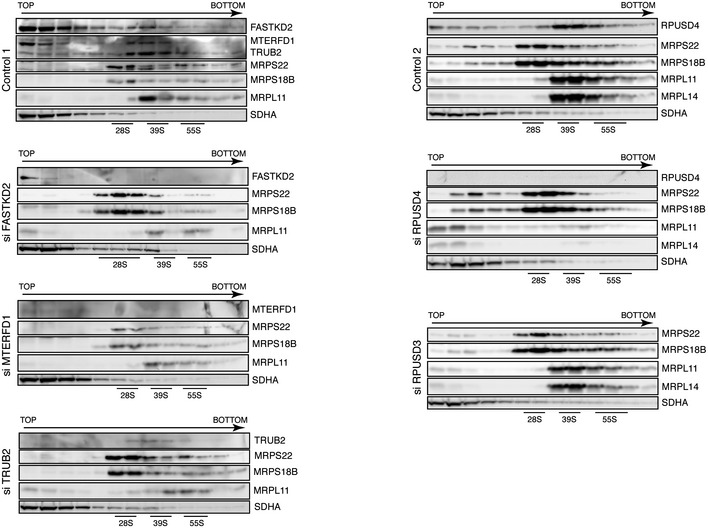
- A, B
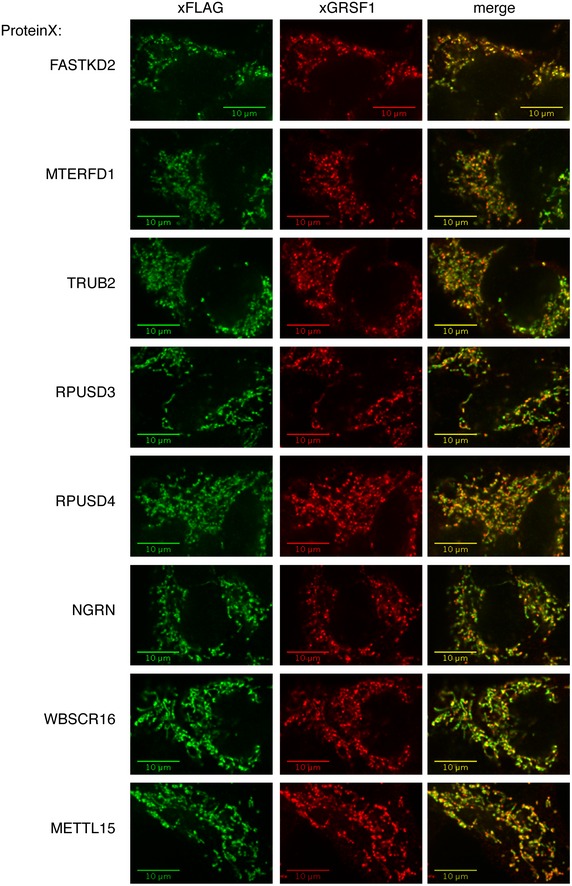
Identification of mitochondrial ribosomal proteins and pseudouridine synthase interacting proteins by sucrose gradient centrifugation in control cells (A) and cells treated with siRNA (B). Individual fractions were separated by SDS–PAGE and immunoblotted with the indicated antibodies. Panel control 2 in (B) is identical to the one shown in (A). The migration of the mt‐SSU (28S), the mt‐LSU (39S), and the mitochondrial monosome (55S) is shown.
- C
Quantification of the levels of the mt‐SSU (28S), the mt‐LSU (39S), and the mitochondrial monosome (55S) normalized to the SDHA levels. The graph represents the relative abundance of individual subunits in cells treated with specified siRNA versus controls. Quantification of the mt‐SSU, mt‐LSU, and monosome was done by averaging the intensity of the signals from three different antibodies directed against specific structural subunits of the ribosome. The bars represent the mean ± SEM.
- D
2D‐immunoblot analysis (BN–PAGE/SDS–PAGE) of pseudouridine synthase interacting proteins. The migration (sizes in kDa) of known protein complexes in the first dimension is indicated on the top of the blot.
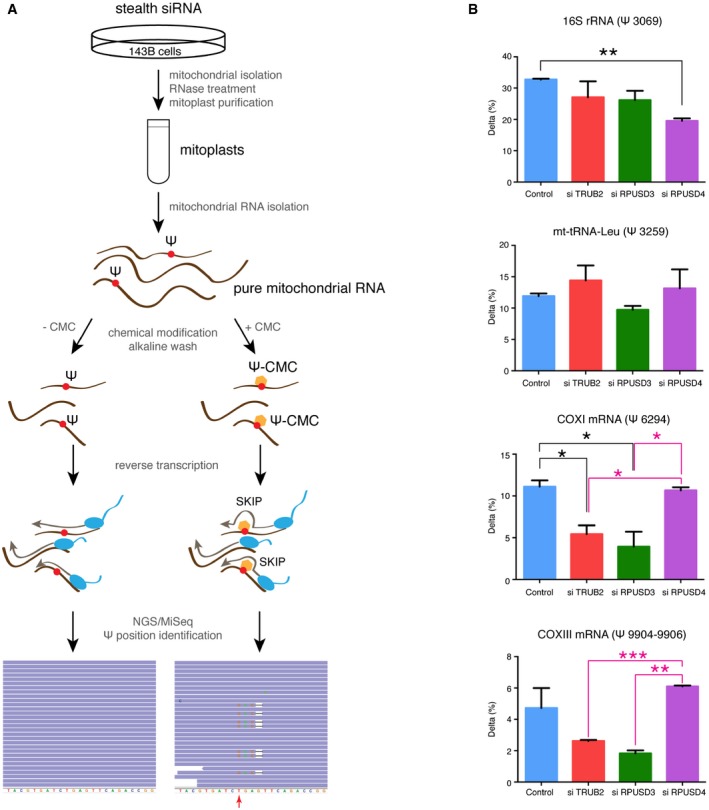
Experimental approach used for the identification of pseudouridine sites in mitochondrial RNA. A combination of the CMC modification and the next‐generation sequencing on pure mitochondrial RNA from control and siRNA‐treated cells was used. CMC‐ψ modification causes skips in reverse transcription rather than stops, resulting in 1‐bp deletions in sequence alignments. The red arrow indicates a putative pseudouridine site.
Quantification of Δ (the difference between the rate of deletions in CMC‐treated versus untreated cells) for individual ψ sites in control and siRNA‐treated cells. The bars represent the mean ± SEM of two independent experiments. P‐values were calculated using a one‐tailed t‐test (*P < 0.05; **P < 0.01; ***P < 0.001).
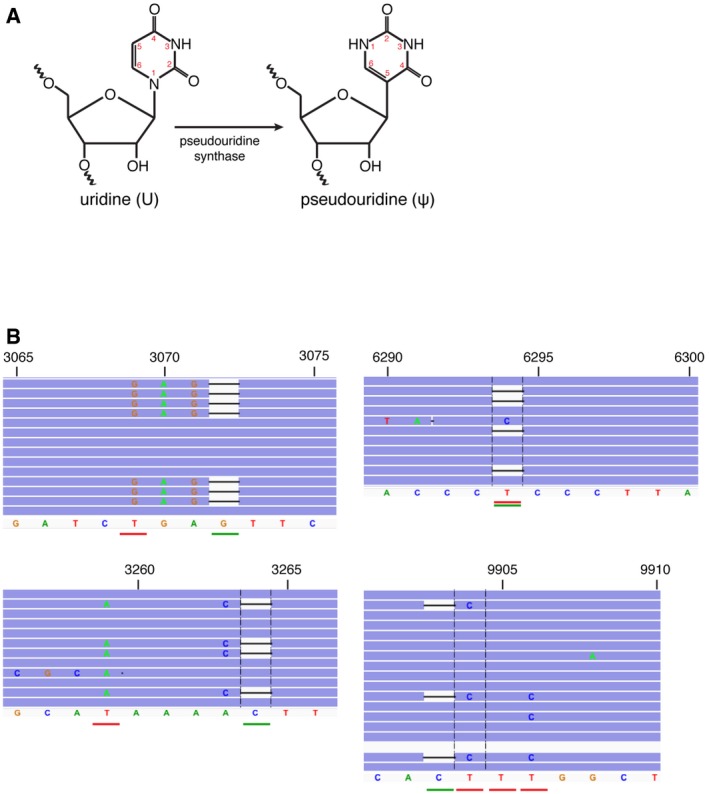
A schematic of an enzymatic reaction of pseudouridine (ψ) synthesis by pseudouridine synthase.
IGV screenshot of reads in CMC‐treated samples mapping to the top four pseudouridylated sites. The position of the deletion is underlined in green, and the putative pseudouridylated site is underlined in red.
- A
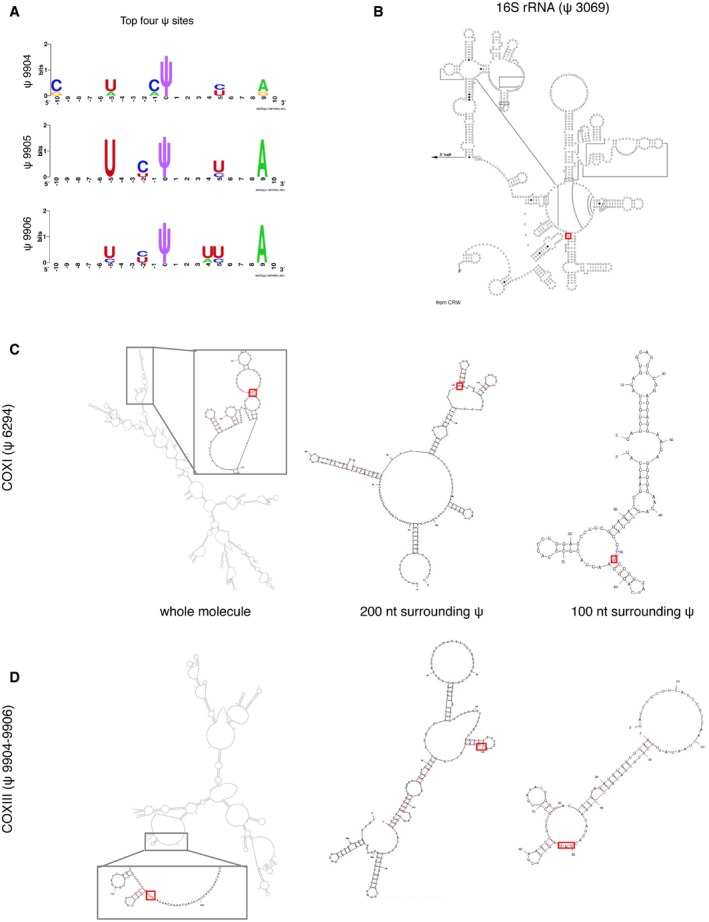
Sequence motifs surrounding the mitochondrial ψ sites generated by WebLogo software at
http://weblogo.berkeley.edu . Three motifs are suggested for the top four ψ sites, because the COXIII (ψ 9904–9906) position is uncertain due to the ambiguity of the sequence alignments. - B–D
Positions of identified ψ sites (red rectangle) within the secondary structures of individual RNAs. (B) Known secondary structure of the 3′‐terminus of 16S rRNA 37 and predicted structures for COXI (C) and COXIII mRNAs (D). Predictions were based either on sequence fragments of 100 nt and 200 nt surrounding each ψ site, or using the whole mRNA molecule as the input. Pseudouridine is predicted to be accessible in all cases, located at a loop or in the boundary between a stem and a loop/bulge structure.
References
-
- Liu N, Pan T (2016) N6‐methyladenosine‐encoded epitranscriptomics. Nat Struct Mol Biol 23: 98–102 - PubMed
-
- Li X, Ma S, Yi C (2016) Pseudouridine: the fifth RNA nucleotide with renewed interests. Curr Opin Chem Biol 33: 108–116 - PubMed
-
- Ofengand J, Bakin A (1997) Mapping to nucleotide resolution of pseudouridine residues in large subunit ribosomal RNAs from representative eukaryotes, prokaryotes, archaebacteria, mitochondria and chloroplasts. J Mol Biol 266: 246–268 - PubMed
-
- Patton JR, Bykhovskaya Y, Mengesha E, Bertolotto C, Fischel‐Ghodsian N (2005) Mitochondrial myopathy and sideroblastic anemia (MLASA): missense mutation in the pseudouridine synthase 1 (PUS1) gene is associated with the loss of tRNA pseudouridylation. J Biol Chem 280: 19823–19828 - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
