

Cyclic AMP Inhibits the Activity and Promotes the Acetylation of Acetyl-CoA Synthetase through Competitive Binding to the ATP/AMP Pocket
- PMID: 27974467
- PMCID: PMC5270480
- DOI: 10.1074/jbc.M116.753640
Cyclic AMP Inhibits the Activity and Promotes the Acetylation of Acetyl-CoA Synthetase through Competitive Binding to the ATP/AMP Pocket
Abstract
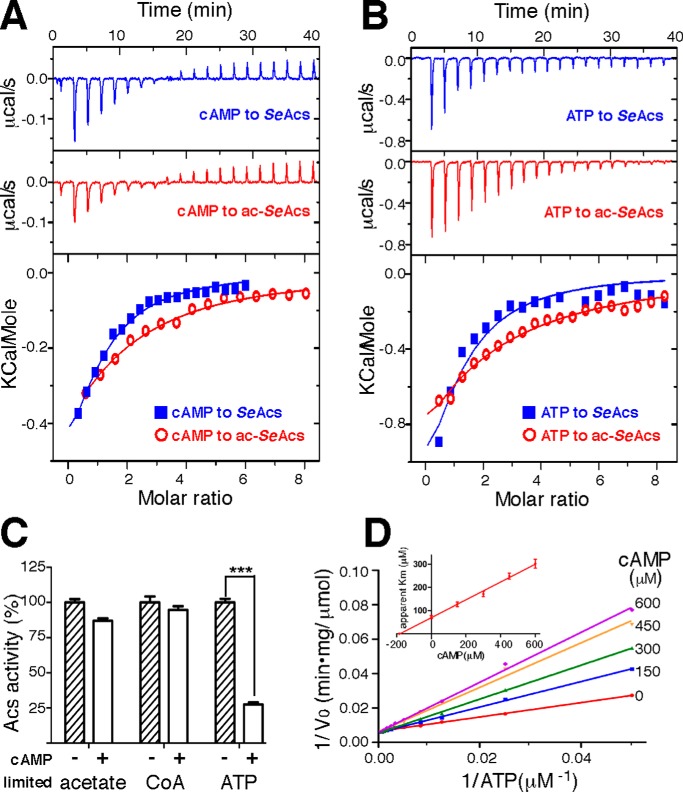
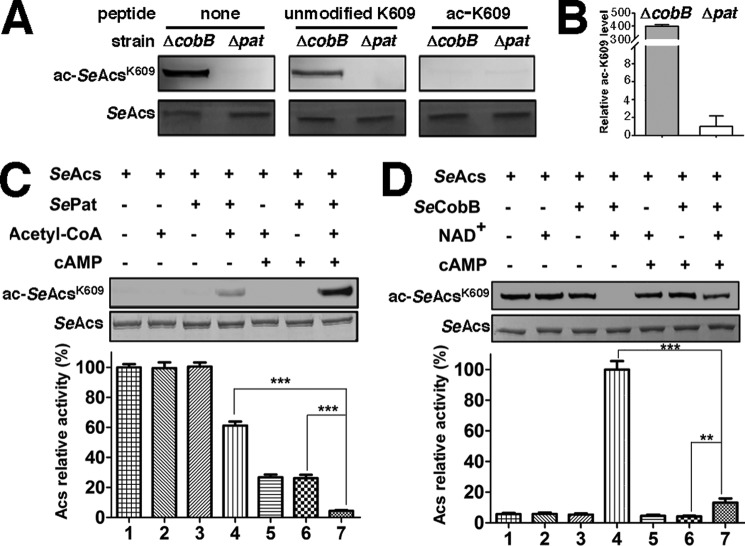
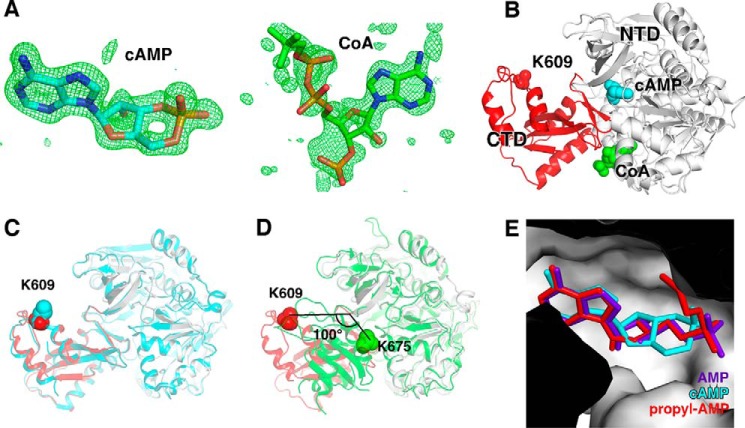
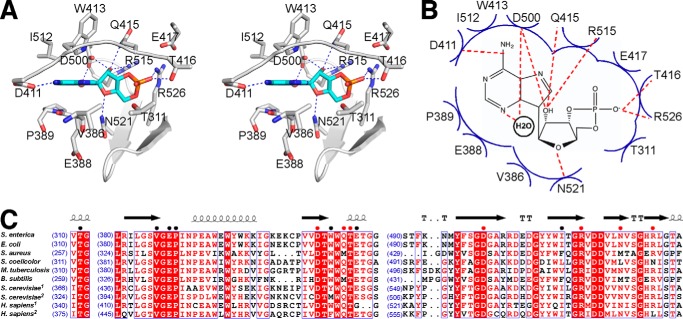
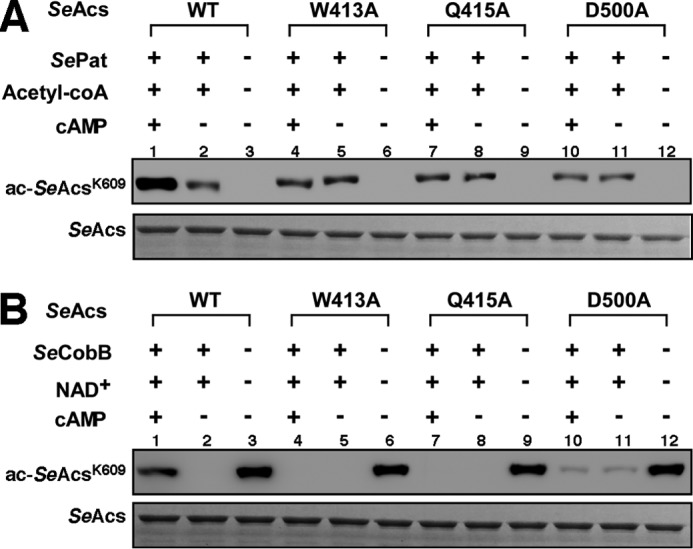
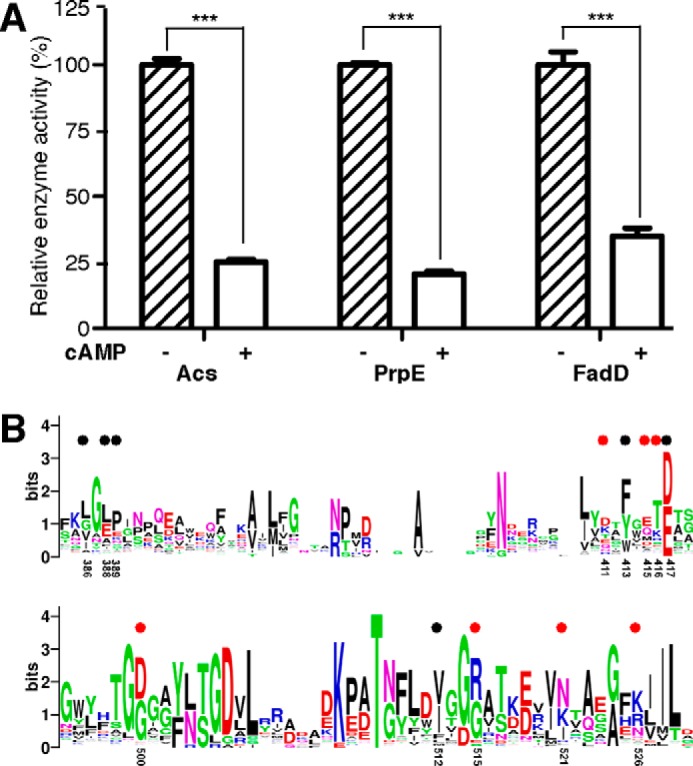
The high-affinity biosynthetic pathway for converting acetate to acetyl-coenzyme A (acetyl-CoA) is catalyzed by the central metabolic enzyme acetyl-coenzyme A synthetase (Acs), which is finely regulated both at the transcriptional level via cyclic AMP (cAMP)-driven trans-activation and at the post-translational level via acetylation inhibition. In this study, we discovered that cAMP directly binds to Salmonella enterica Acs (SeAcs) and inhibits its activity in a substrate-competitive manner. In addition, cAMP binding increases SeAcs acetylation by simultaneously promoting Pat-dependent acetylation and inhibiting CobB-dependent deacetylation, resulting in enhanced SeAcs inhibition. A crystal structure study and site-directed mutagenesis analyses confirmed that cAMP binds to the ATP/AMP pocket of SeAcs, and restrains SeAcs in an open conformation. The cAMP contact residues are well conserved from prokaryotes to eukaryotes, suggesting a general regulatory mechanism of cAMP on Acs.
Keywords: acetyl-CoA synthetase; acetylation; crystal structure; cyclic AMP (cAMP); post-translational modification (PTM).
© 2017 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Pietrocola F., Galluzzi L., Bravo-San Pedro J. M., Madeo F., and Kroemer G. (2015) Acetyl coenzyme A: a central metabolite and second messenger. Cell Metab. 21, 805–821 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
