

Bacterial Proteasomes: Mechanistic and Functional Insights
- PMID: 27974513
- PMCID: PMC5312241
- DOI: 10.1128/MMBR.00036-16
Bacterial Proteasomes: Mechanistic and Functional Insights
Abstract
Regulated proteolysis is essential for the normal physiology of all organisms. While all eukaryotes and archaea use proteasomes for protein degradation, only certain orders of bacteria have proteasomes, whose functions are likely as diverse as the species that use them. In this review, we discuss the most recent developments in the understanding of how proteins are targeted to proteasomes for degradation, including ATP-dependent and -independent mechanisms, and the roles of proteasome-dependent degradation in protein quality control and the regulation of cellular physiology. Furthermore, we explore newly established functions of proteasome system accessory factors that function independently of proteolysis.
Keywords: proteasome.
Copyright © 2016 American Society for Microbiology.
Figures
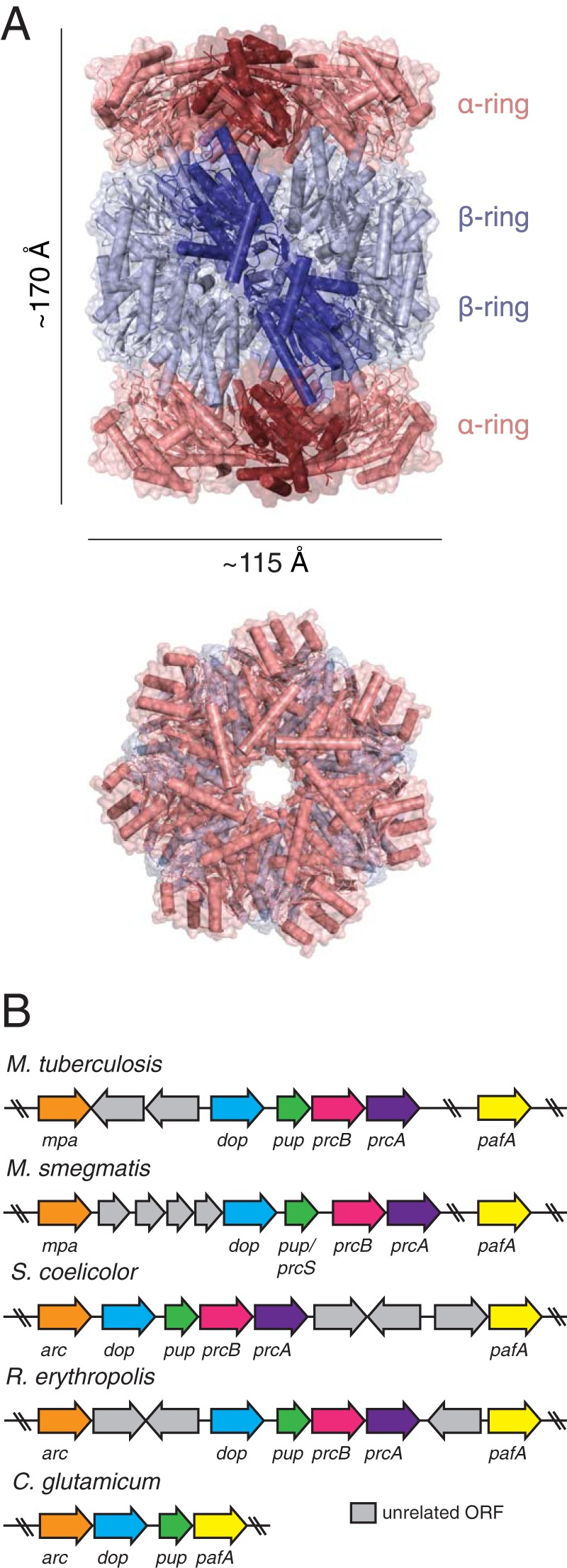
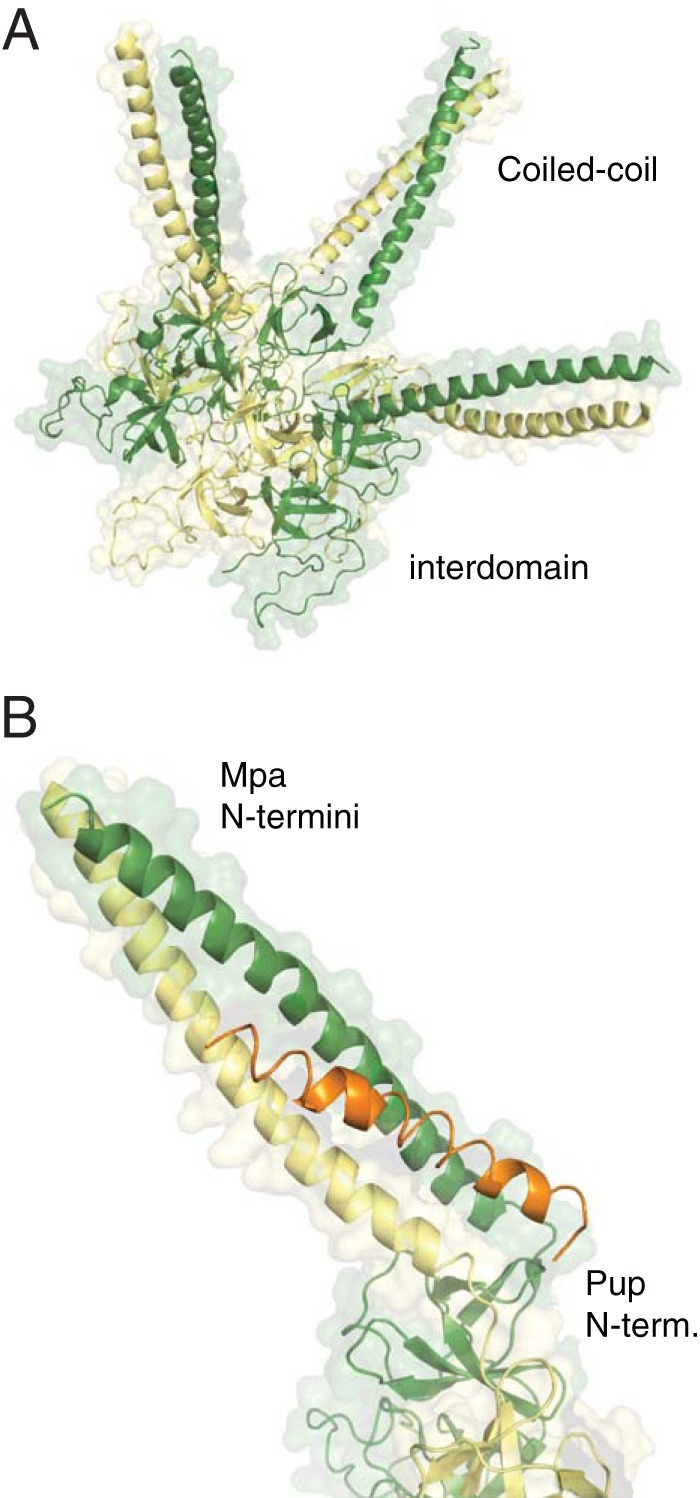
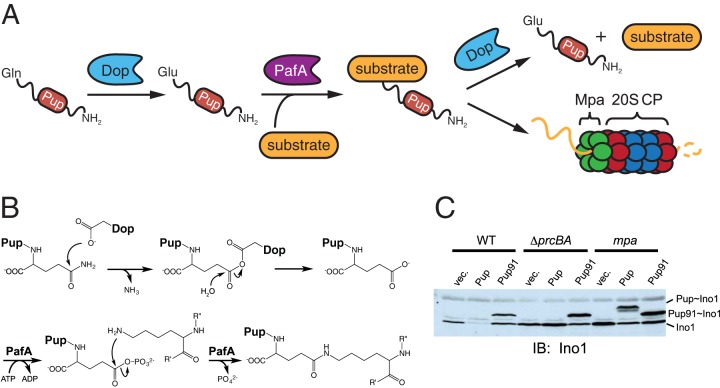

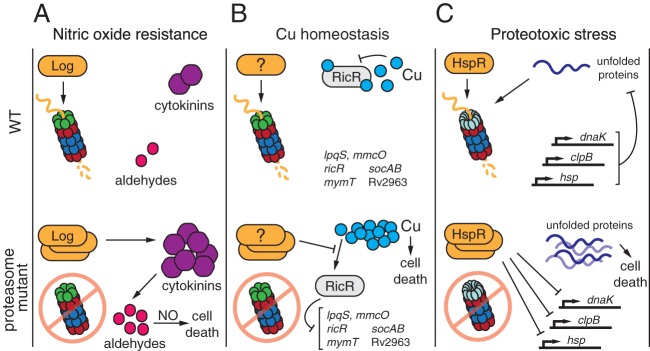
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
