

Can invertebrates see the e-vector of polarization as a separate modality of light?
- PMID: 27974532
- PMCID: PMC5201003
- DOI: 10.1242/jeb.139899
Can invertebrates see the e-vector of polarization as a separate modality of light?
Abstract
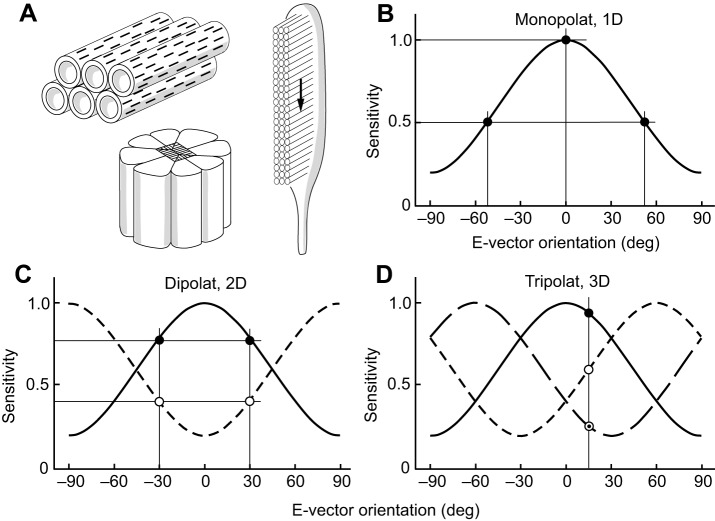
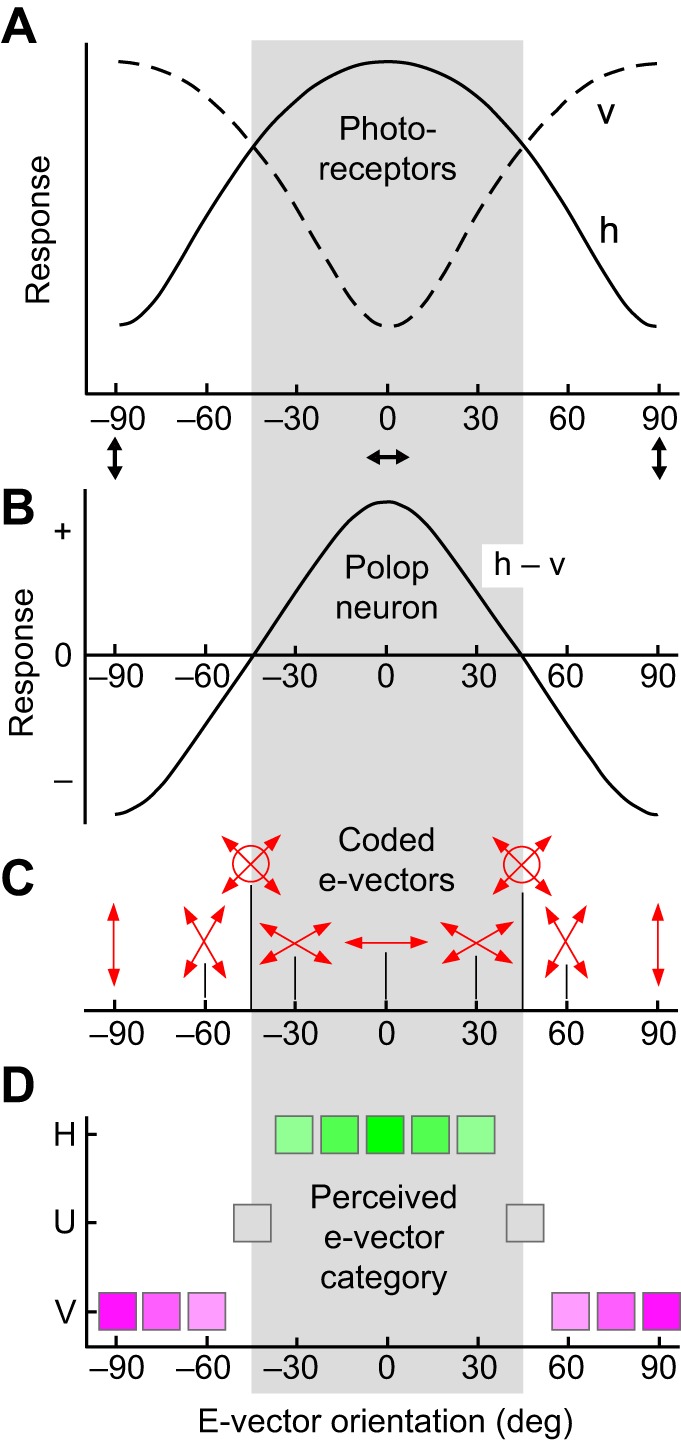
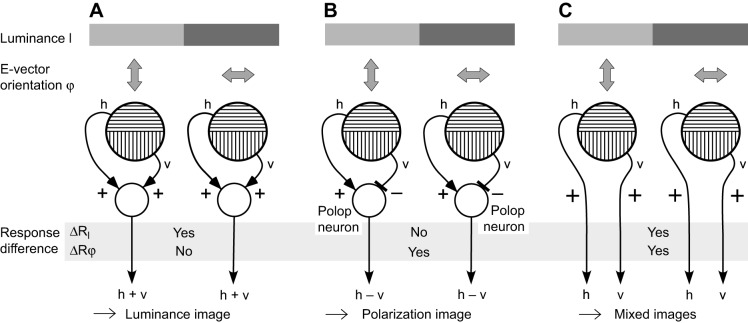
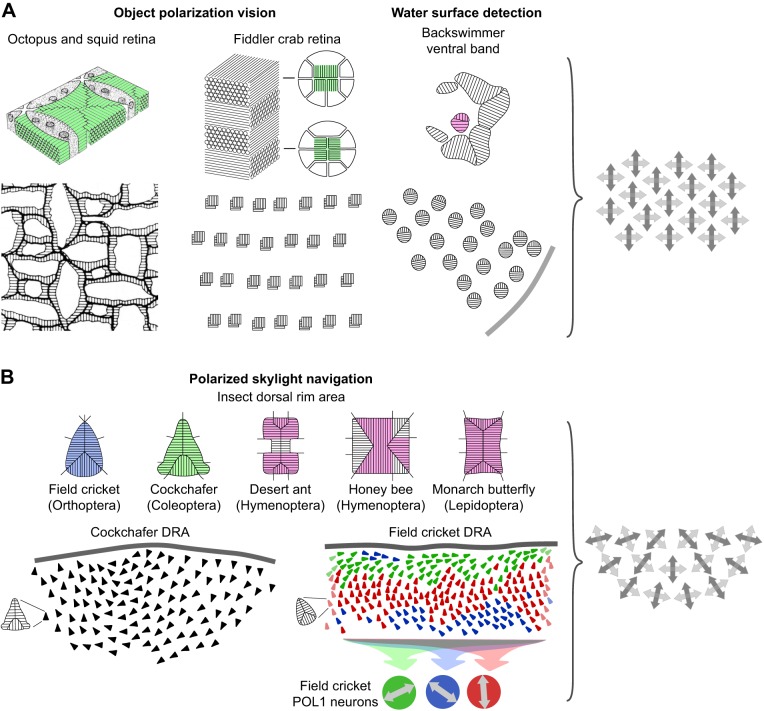
The visual world is rich in linearly polarized light stimuli, which are hidden from the human eye. But many invertebrate species make use of polarized light as a source of valuable visual information. However, exploiting light polarization does not necessarily imply that the electric (e)-vector orientation of polarized light can be perceived as a separate modality of light. In this Review, I address the question of whether invertebrates can detect specific e-vector orientations in a manner similar to that of humans perceiving spectral stimuli as specific hues. To analyze e-vector orientation, the signals of at least three polarization-sensitive sensors (analyzer channels) with different e-vector tuning axes must be compared. The object-based, imaging polarization vision systems of cephalopods and crustaceans, as well as the water-surface detectors of flying backswimmers, use just two analyzer channels. Although this excludes the perception of specific e-vector orientations, a two-channel system does provide a coarse, categoric analysis of polarized light stimuli, comparable to the limited color sense of dichromatic, 'color-blind' humans. The celestial compass of insects employs three or more analyzer channels. However, that compass is multimodal, i.e. e-vector information merges with directional information from other celestial cues, such as the solar azimuth and the spectral gradient in the sky, masking e-vector information. It seems that invertebrate organisms take no interest in the polarization details of visual stimuli, but polarization vision grants more practical benefits, such as improved object detection and visual communication for cephalopods and crustaceans, compass readings to traveling insects, or the alert 'water below!' to water-seeking bugs.
Keywords: Celestial compass; E-vector perception; Invertebrates; Polarization imaging; Polarization vision; Polarization-opponent neurons.
© 2016. Published by The Company of Biologists Ltd.
Conflict of interest statement
The author declares no competing or financial interests.
Figures

References
-
- Brunner D. and Labhart T. (1987). Behavioural evidence for polarization vision in crickets. Physiol. Entomol. 12, 1-10. 10.1111/j.1365-3032.1987.tb00718.x - DOI
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
