

On the Spatial Organization of mRNA, Plasmids, and Ribosomes in a Bacterial Host Overexpressing Membrane Proteins
- PMID: 27977669
- PMCID: PMC5201305
- DOI: 10.1371/journal.pgen.1006523
On the Spatial Organization of mRNA, Plasmids, and Ribosomes in a Bacterial Host Overexpressing Membrane Proteins
Abstract
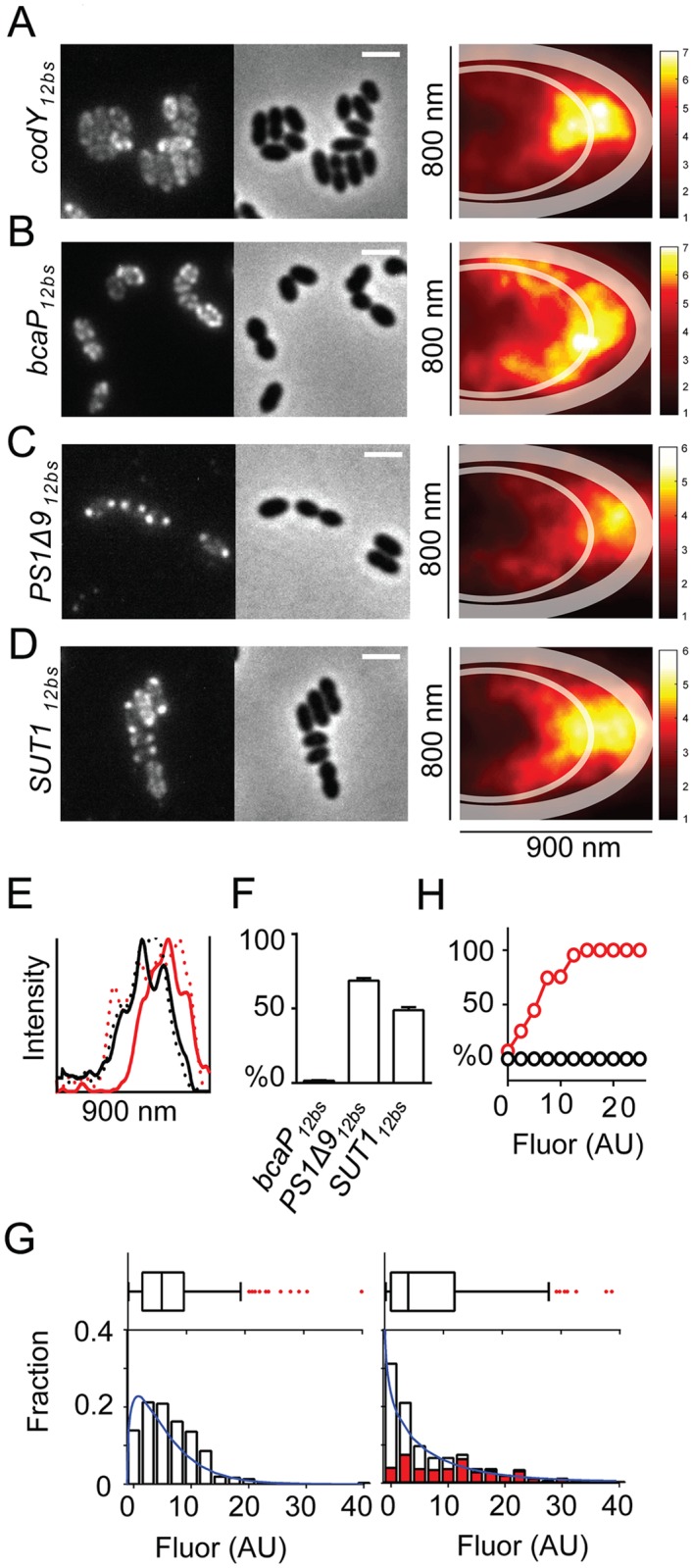
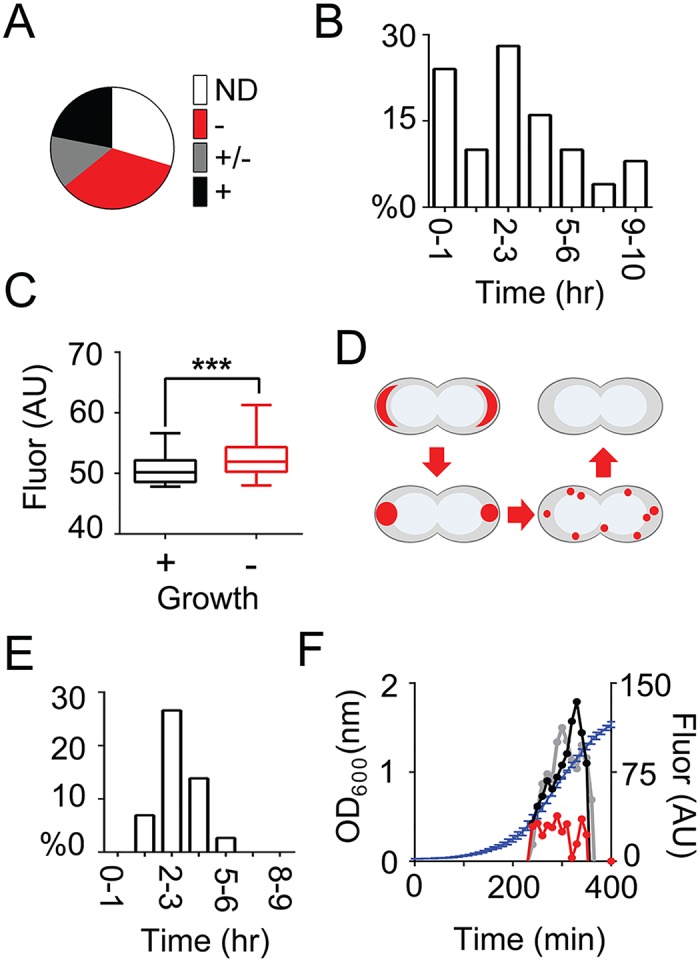
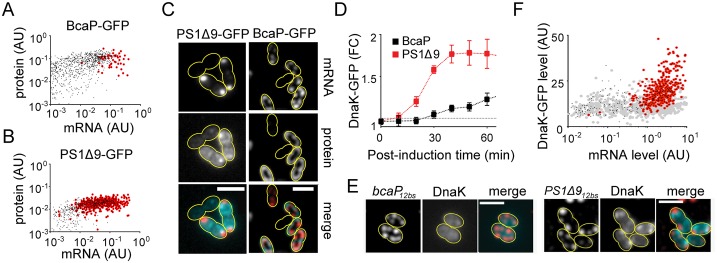
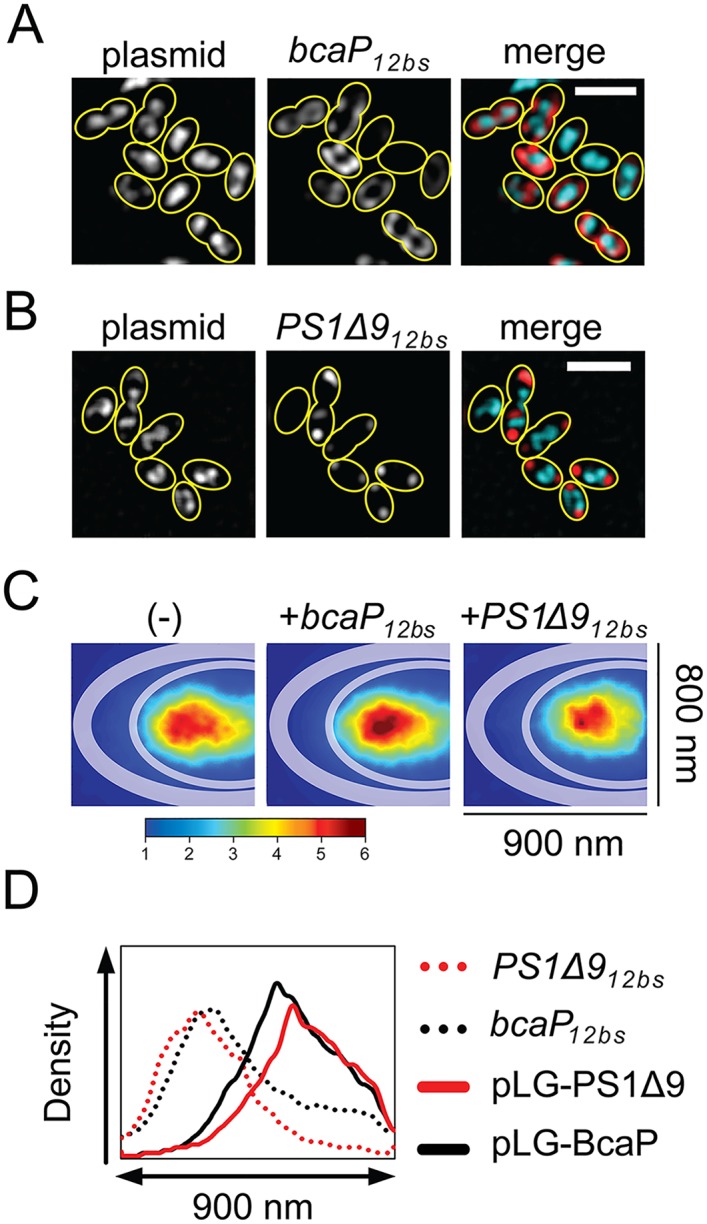
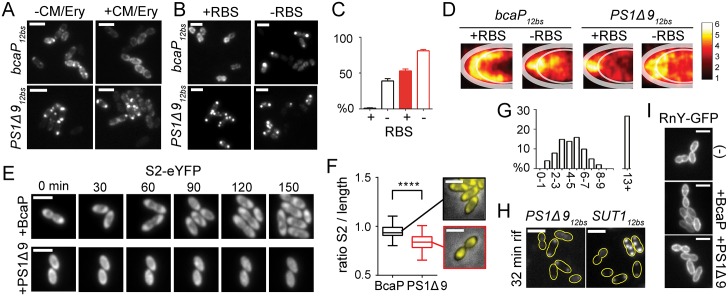
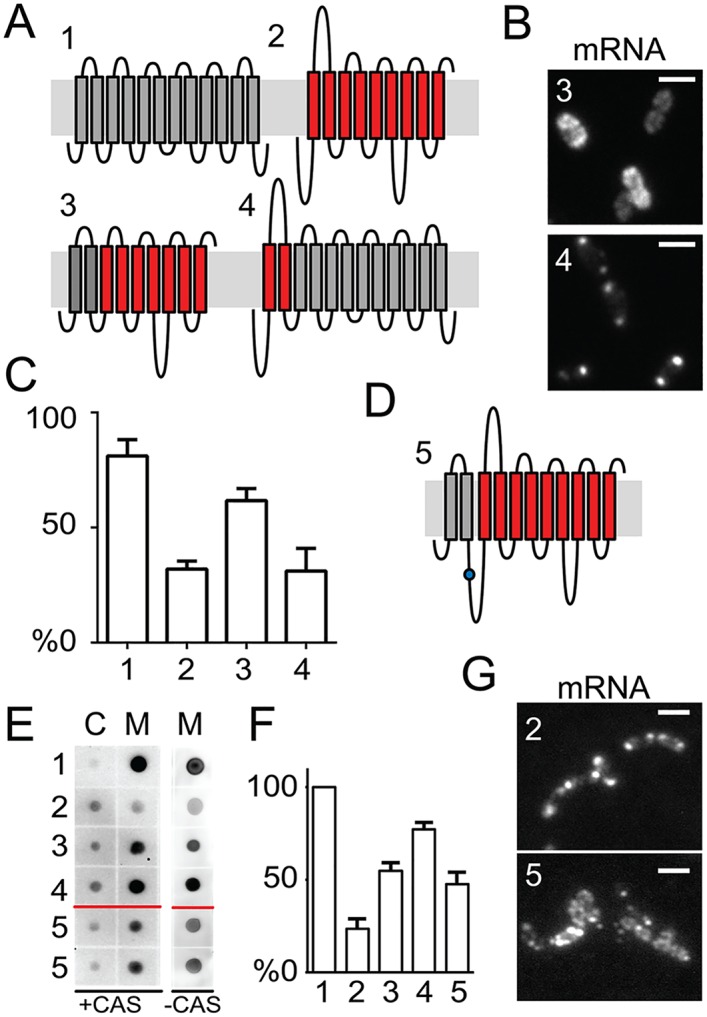
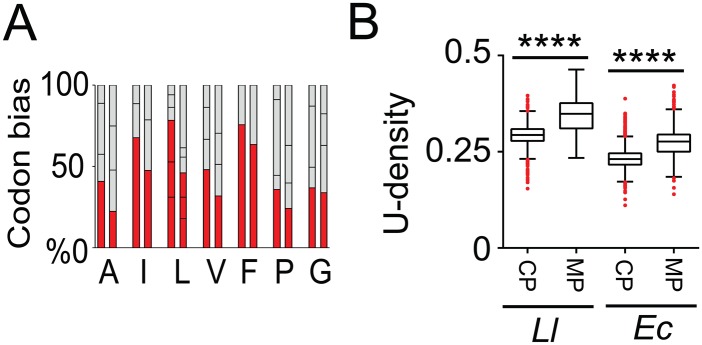
By using fluorescence imaging, we provide a time-resolved single-cell view on coupled defects in transcription, translation, and growth during expression of heterologous membrane proteins in Lactococcus lactis. Transcripts encoding poorly produced membrane proteins accumulate in mRNA-dense bodies at the cell poles, whereas transcripts of a well-expressed homologous membrane protein show membrane-proximal localization in a translation-dependent fashion. The presence of the aberrant polar mRNA foci correlates with cessation of cell division, which is restored once these bodies are cleared. In addition, activation of the heat-shock response and a loss of nucleoid-occluded ribosomes are observed. We show that the presence of a native-like N-terminal domain is key to SRP-dependent membrane localization and successful production of membrane proteins. The work presented gives new insights and detailed understanding of aberrant membrane protein biogenesis, which can be used for strategies to optimize membrane protein production.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Development of an inducible system to control and easily monitor gene expression in Lactococcus lactis.Plasmid. 2004 May;51(3):256-64. doi: 10.1016/j.plasmid.2004.02.004. Plasmid. 2004. PMID: 15109832
-
recA gene involvement in oxidative and thermal stress in Lactococcus lactis.Dev Biol Stand. 1995;85:455-67. Dev Biol Stand. 1995. PMID: 8586217
-
Spatial organization of bacterial transcription and translation.Proc Natl Acad Sci U S A. 2016 Aug 16;113(33):9286-91. doi: 10.1073/pnas.1604995113. Epub 2016 Aug 2. Proc Natl Acad Sci U S A. 2016. PMID: 27486246 Free PMC article.
-
The Evolution of gene regulation research in Lactococcus lactis.FEMS Microbiol Rev. 2017 Aug 1;41(Supp_1):S220-S243. doi: 10.1093/femsre/fux028. FEMS Microbiol Rev. 2017. PMID: 28830093 Review.
-
Examination of post-transcriptional regulations in prokaryotes by integrative biology.C R Biol. 2009 Nov;332(11):958-73. doi: 10.1016/j.crvi.2009.09.005. Epub 2009 Oct 14. C R Biol. 2009. PMID: 19909919 Review.
Cited by
-
Real-Time Messenger RNA Dynamics in Bacillus subtilis.Front Microbiol. 2021 Nov 18;12:760857. doi: 10.3389/fmicb.2021.760857. eCollection 2021. Front Microbiol. 2021. PMID: 34867890 Free PMC article.
-
Nucleoid Size Scaling and Intracellular Organization of Translation across Bacteria.Cell. 2019 May 30;177(6):1632-1648.e20. doi: 10.1016/j.cell.2019.05.017. Cell. 2019. PMID: 31150626 Free PMC article.
-
mRNA targeting eliminates the need for the signal recognition particle during membrane protein insertion in bacteria.Cell Rep. 2023 Mar 28;42(3):112140. doi: 10.1016/j.celrep.2023.112140. Epub 2023 Feb 25. Cell Rep. 2023. PMID: 36842086 Free PMC article.
-
Illuminating Messengers: An Update and Outlook on RNA Visualization in Bacteria.Front Microbiol. 2017 Jun 22;8:1161. doi: 10.3389/fmicb.2017.01161. eCollection 2017. Front Microbiol. 2017. PMID: 28690601 Free PMC article. Review.
-
Macromolecular Crowding, Phase Separation, and Homeostasis in the Orchestration of Bacterial Cellular Functions.Chem Rev. 2024 Feb 28;124(4):1899-1949. doi: 10.1021/acs.chemrev.3c00622. Epub 2024 Feb 8. Chem Rev. 2024. PMID: 38331392 Free PMC article. Review.
References
-
- Kunji ERS, Slotboom D-J, Poolman B. Lactococcus lactis as host for overproduction of functional membrane proteins. Biochim Biophys Acta. 2003;1610: 97–108. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
