

LGR4 Is a Direct Target of MicroRNA-34a and Modulates the Proliferation and Migration of Retinal Pigment Epithelial ARPE-19 Cells
- PMID: 27977785
- PMCID: PMC5158047
- DOI: 10.1371/journal.pone.0168320
LGR4 Is a Direct Target of MicroRNA-34a and Modulates the Proliferation and Migration of Retinal Pigment Epithelial ARPE-19 Cells
Abstract
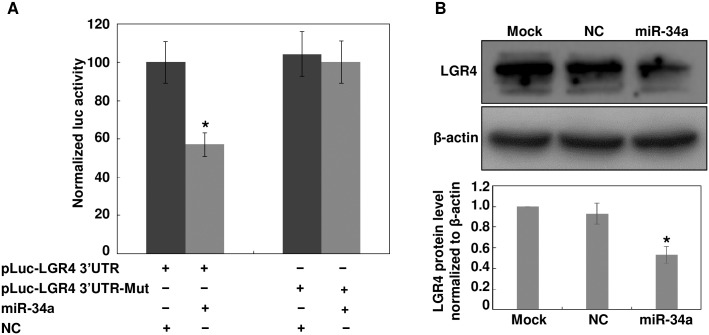
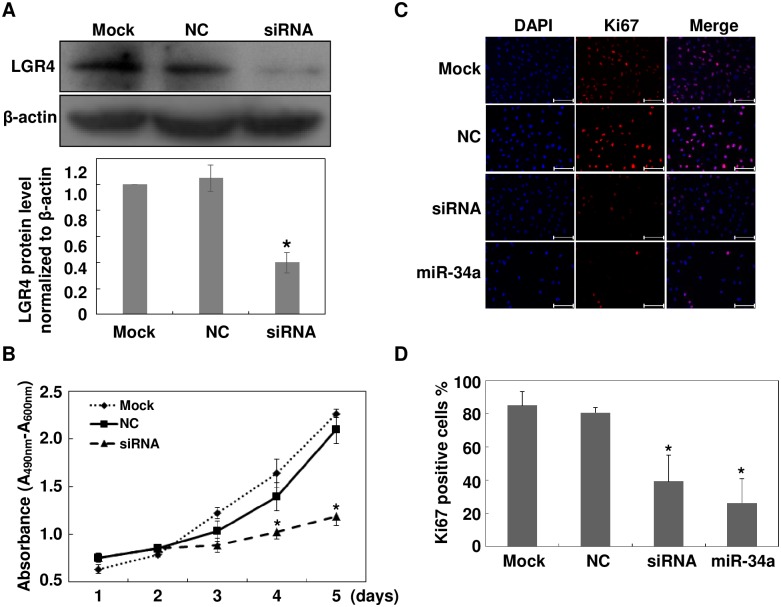
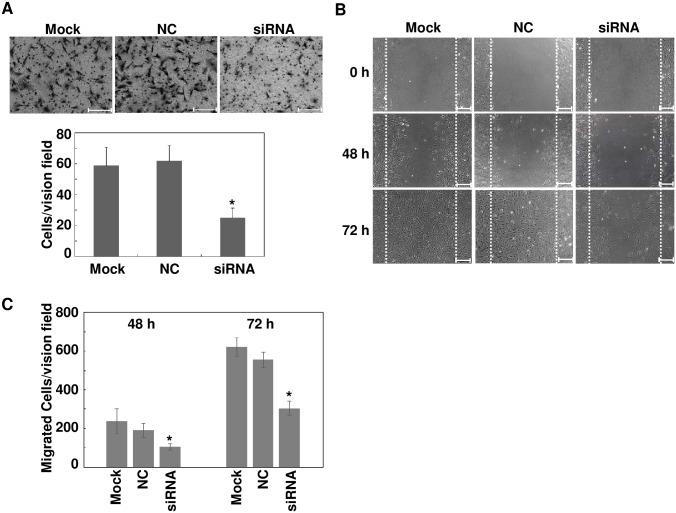
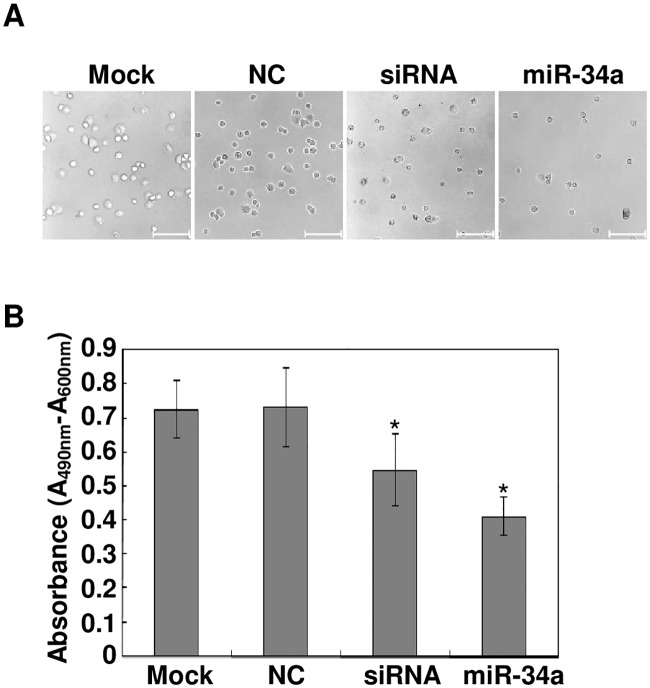
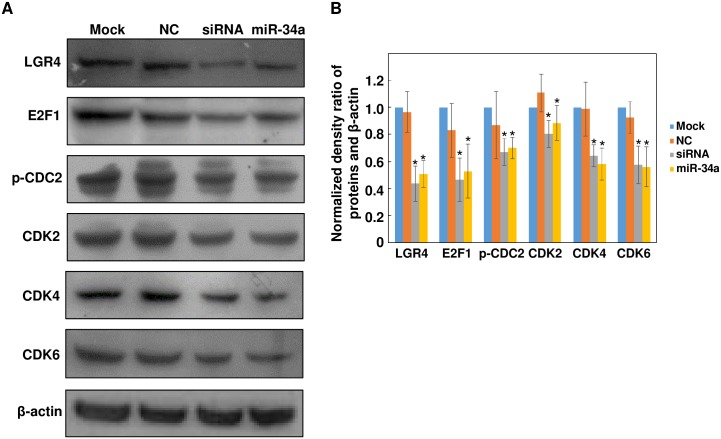
The pathology of proliferative vitreoretinopathy and proliferative diabetic retinopathy is linked to proliferation, migration, and adhesion of the retinal pigment epithelium. MicroRNA-34a (miR-34a) expression modulates changes in proliferation and migration of retinal pigment epithelial cell line ARPE-19. In this study, we determined that miR-34a interacts with LGR4, identified by bioinformatics using TargetScan Human 5.0, to affect these changes. Double luciferase gene reporter assay confirmed miR-34a involvement in mediating control. miR-34a mimic transfection decreased LGR4 expression. Western blot analysis documented corresponding protein expression inhibition. MTS, Ki67 immunostaining, scratch and transwell testing, along with attachment assay showed that miR-34a upregulation inhibited ARPE-19 cell proliferation, migration and attachment partly through downregulation of LGR4 protein expression. Western blot analysis revealed that both miR-34a upregulation and LGR4 downregulation induced declines in E2F1, p-CDC2, CDK2, CDK4 and CDK6 protein expression. Taken together, miR-34a gene expression upregulation inhibits ARPE-19 cell proliferation, migration and adhesion partly by suppressing LGR4 expression. These results substantiate earlier indications that both miR-34a and LGR4 are potential drug targets to prevent fibrosis in a clinical setting.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Regulation of Ras homolog family member G by microRNA-124 regulates proliferation and migration of human retinal pigment epithelial cells.Sci Rep. 2020 Sep 22;10(1):15420. doi: 10.1038/s41598-020-72360-5. Sci Rep. 2020. PMID: 32963317 Free PMC article.
-
Inhibitory effect of microRNA-34a on retinal pigment epithelial cell proliferation and migration.Invest Ophthalmol Vis Sci. 2013 Oct 3;54(10):6481-8. doi: 10.1167/iovs.13-11873. Invest Ophthalmol Vis Sci. 2013. PMID: 23970470
-
The Interplay of MicroRNA-34a, LGR4, EMT-Associated Factors, and MMP2 in Regulating Uveal Melanoma Cells.Invest Ophthalmol Vis Sci. 2019 Oct 1;60(13):4503-4510. doi: 10.1167/iovs.18-26477. Invest Ophthalmol Vis Sci. 2019. PMID: 31661551
-
MicroRNA-182 Suppresses HGF/SF-Induced Increases in Retinal Pigment Epithelial Cell Proliferation and Migration through Targeting c-Met.PLoS One. 2016 Dec 9;11(12):e0167684. doi: 10.1371/journal.pone.0167684. eCollection 2016. PLoS One. 2016. PMID: 27936052 Free PMC article.
-
MicroRNA-34a inhibits uveal melanoma cell proliferation and migration through downregulation of c-Met.Invest Ophthalmol Vis Sci. 2009 Apr;50(4):1559-65. doi: 10.1167/iovs.08-2681. Epub 2008 Nov 21. Invest Ophthalmol Vis Sci. 2009. PMID: 19029026
Cited by
-
The HIF-1α/p53/miRNA-34a/Klotho axis in retinal pigment epithelial cells promotes subretinal fibrosis and exacerbates choroidal neovascularization.J Cell Mol Med. 2021 Feb;25(3):1700-1711. doi: 10.1111/jcmm.16272. Epub 2021 Jan 12. J Cell Mol Med. 2021. PMID: 33438362 Free PMC article.
-
Regulation of Ras homolog family member G by microRNA-124 regulates proliferation and migration of human retinal pigment epithelial cells.Sci Rep. 2020 Sep 22;10(1):15420. doi: 10.1038/s41598-020-72360-5. Sci Rep. 2020. PMID: 32963317 Free PMC article.
-
MicroRNAs in the Vitreous Humor of Patients with Retinal Detachment and a Different Grading of Proliferative Vitreoretinopathy: A Pilot Study.Transl Vis Sci Technol. 2020 May 22;9(6):23. doi: 10.1167/tvst.9.6.23. eCollection 2020 May. Transl Vis Sci Technol. 2020. PMID: 32821520 Free PMC article.
-
miRNA-124 regulates palmitic acid-induced epithelial-mesenchymal transition and cell migration in human retinal pigment epithelial cells by targeting LIN7C.Exp Ther Med. 2022 Jun 1;24(1):481. doi: 10.3892/etm.2022.11408. eCollection 2022 Jul. Exp Ther Med. 2022. PMID: 35761801 Free PMC article.
-
Biological Involvement of MicroRNAs in Proliferative Vitreoretinopathy.Transl Vis Sci Technol. 2017 Jul 10;6(4):5. doi: 10.1167/tvst.6.4.5. eCollection 2017 Jul. Transl Vis Sci Technol. 2017. PMID: 28706757 Free PMC article. Review.
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
