

Targeting amino acid metabolism for cancer therapy
- PMID: 27988359
- PMCID: PMC5429979
- DOI: 10.1016/j.drudis.2016.12.003
Targeting amino acid metabolism for cancer therapy
Abstract
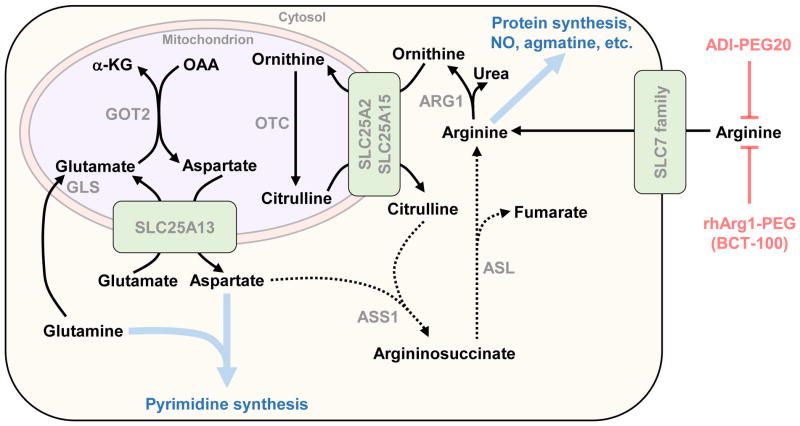
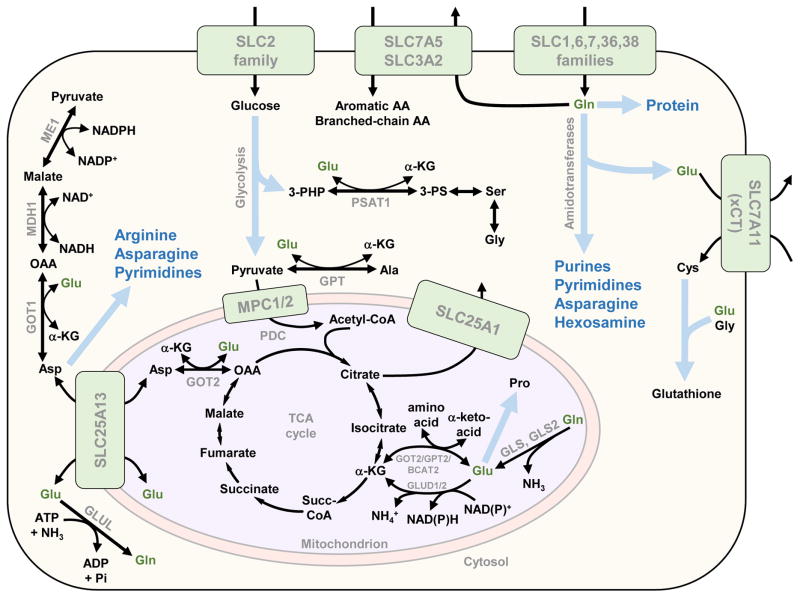
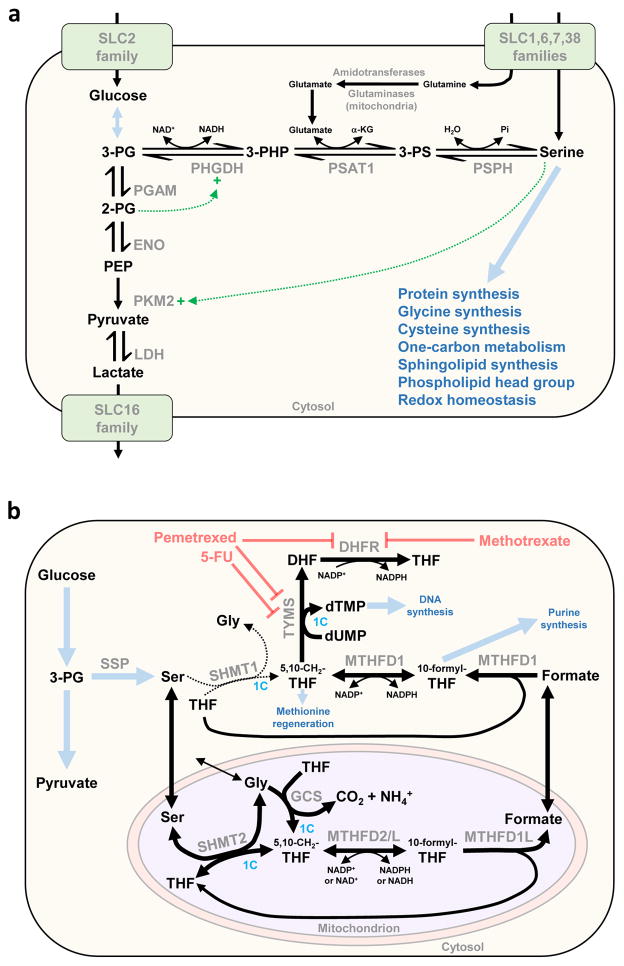
To support sustained biomass accumulation, tumor cells undergo metabolic reprogramming. Nutrient transporters and metabolic enzymes are regulated by the same oncogenic signals that drive cell-cycle progression. Some of the earliest cancer therapies used antimetabolites to disrupt tumor metabolism, and there is now renewed interest in developing drugs that target metabolic dependencies. Many cancers exhibit increased demand for specific amino acids, and become dependent on either an exogenous supply or upregulated de novo synthesis. Strategies to exploit such 'metabolic addictions' include depleting amino acids in blood serum, blocking uptake by transporters and inhibiting biosynthetic or catabolic enzymes. Recent findings highlight the importance of using appropriate model systems and identifying target patient groups as potential therapies advance into the clinic.
Copyright © 2016 Elsevier Ltd. All rights reserved.
Conflict of interest statement
The authors have no conflicts of interest to declare.
Figures
References
-
- Eagle H. Nutrition needs of mammalian cells in tissue culture. Science. 1955;122:501–514. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
