

Chromatin Constrains the Initiation and Elongation of DNA Replication
- PMID: 27989437
- PMCID: PMC5256687
- DOI: 10.1016/j.molcel.2016.10.035
Chromatin Constrains the Initiation and Elongation of DNA Replication
Abstract
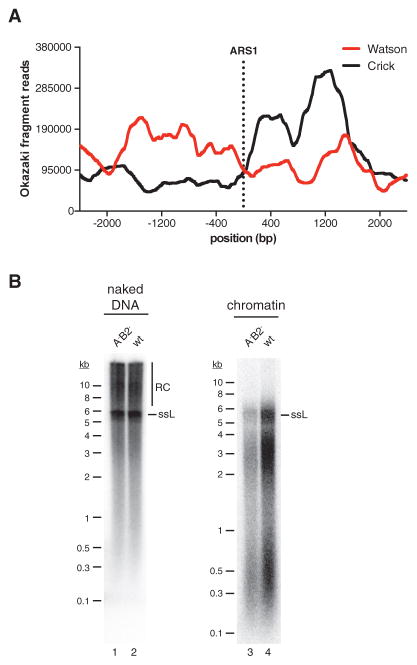
Eukaryotic chromosomal DNA is faithfully replicated in a complex series of cell-cycle-regulated events that are incompletely understood. Here we report the reconstitution of DNA replication free in solution with purified proteins from the budding yeast Saccharomyces cerevisiae. The system recapitulates regulated bidirectional origin activation; synthesis of leading and lagging strands by the three replicative DNA polymerases Pol α, Pol δ, and Pol ε; and canonical maturation of Okazaki fragments into continuous daughter strands. We uncover a dual regulatory role for chromatin during DNA replication: promoting origin dependence and determining Okazaki fragment length by restricting Pol δ progression. This system thus provides a functional platform for the detailed mechanistic analysis of eukaryotic chromosome replication.
Keywords: DNA polymerase; DNA replication; McM2-7; ORC; Okazaki fragment; chromatin; lagging strand; leading strand; replication origin.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures

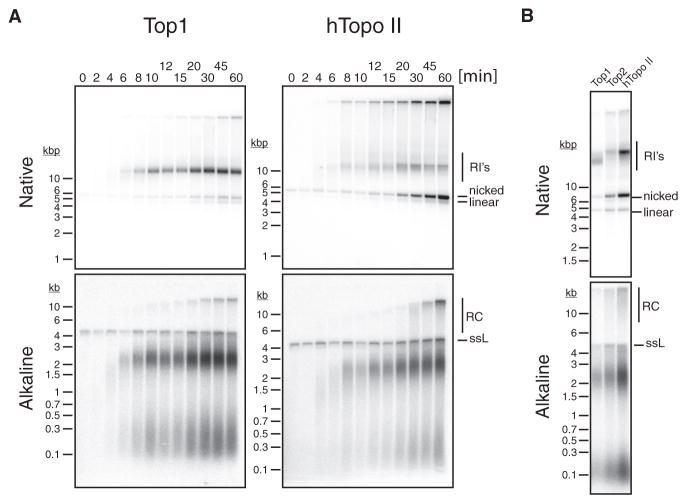
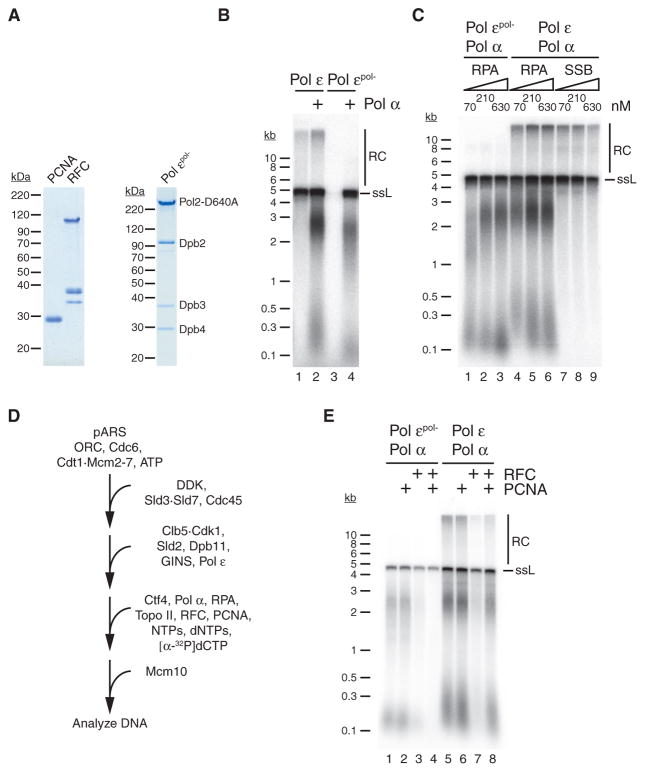

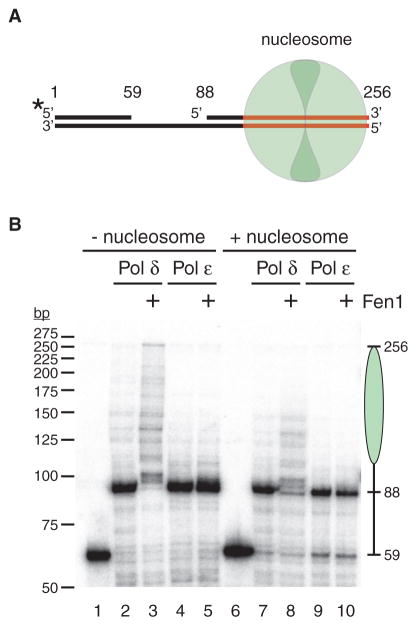
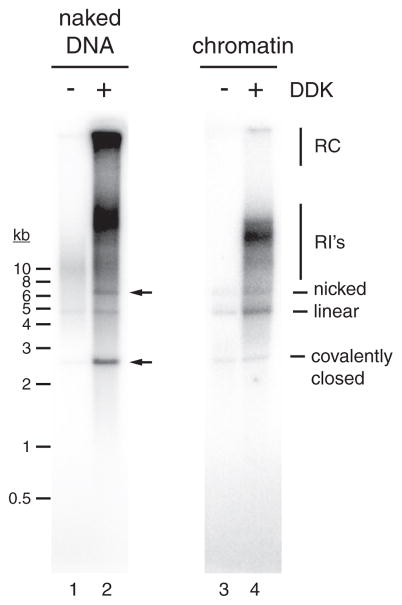
References
-
- Alabert C, Groth A. Chromatin replication and epigenome maintenance. Nat Rev Mol Cell Biol. 2012;13:153–167. - PubMed
-
- Ayyagari R, Gomes XV, Gordenin DA, Burgers PM. Okazaki fragment maturation in yeast. I. Distribution of functions between FEN1 AND DNA2. J Biol Chem. 2003;278:1618–1625. - PubMed
-
- Clément C, Almouzni G. MCM2 binding to histones H3–H4 and ASF1 supports a tetramer-to-dimer model for histone inheritance at the replication fork. Nat Struct Mol Biol. 2015;22:587–589. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
