

Chromatin Controls DNA Replication Origin Selection, Lagging-Strand Synthesis, and Replication Fork Rates
- PMID: 27989438
- PMCID: PMC5222724
- DOI: 10.1016/j.molcel.2016.11.016
Chromatin Controls DNA Replication Origin Selection, Lagging-Strand Synthesis, and Replication Fork Rates
Abstract
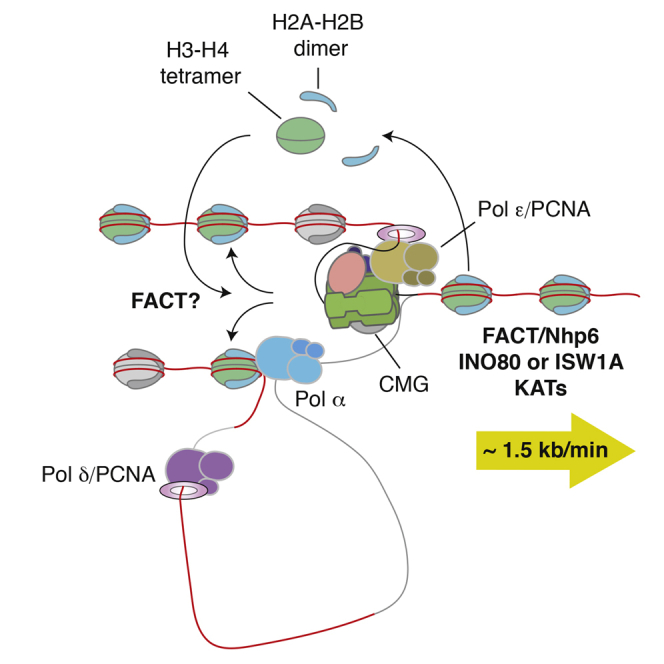
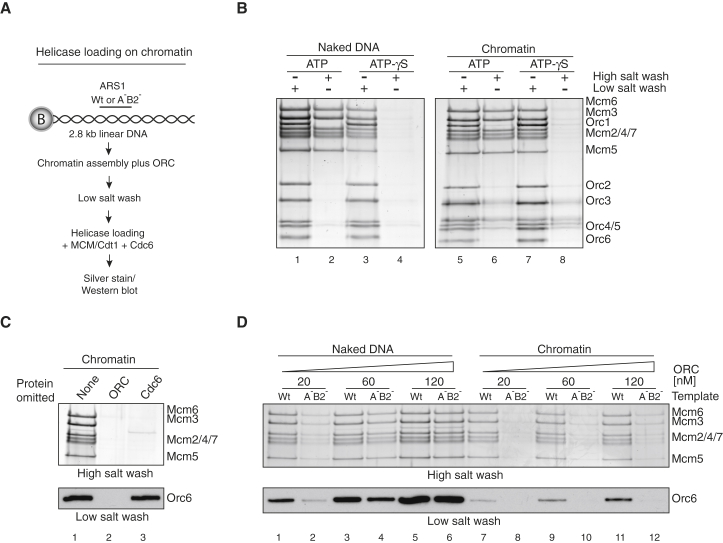
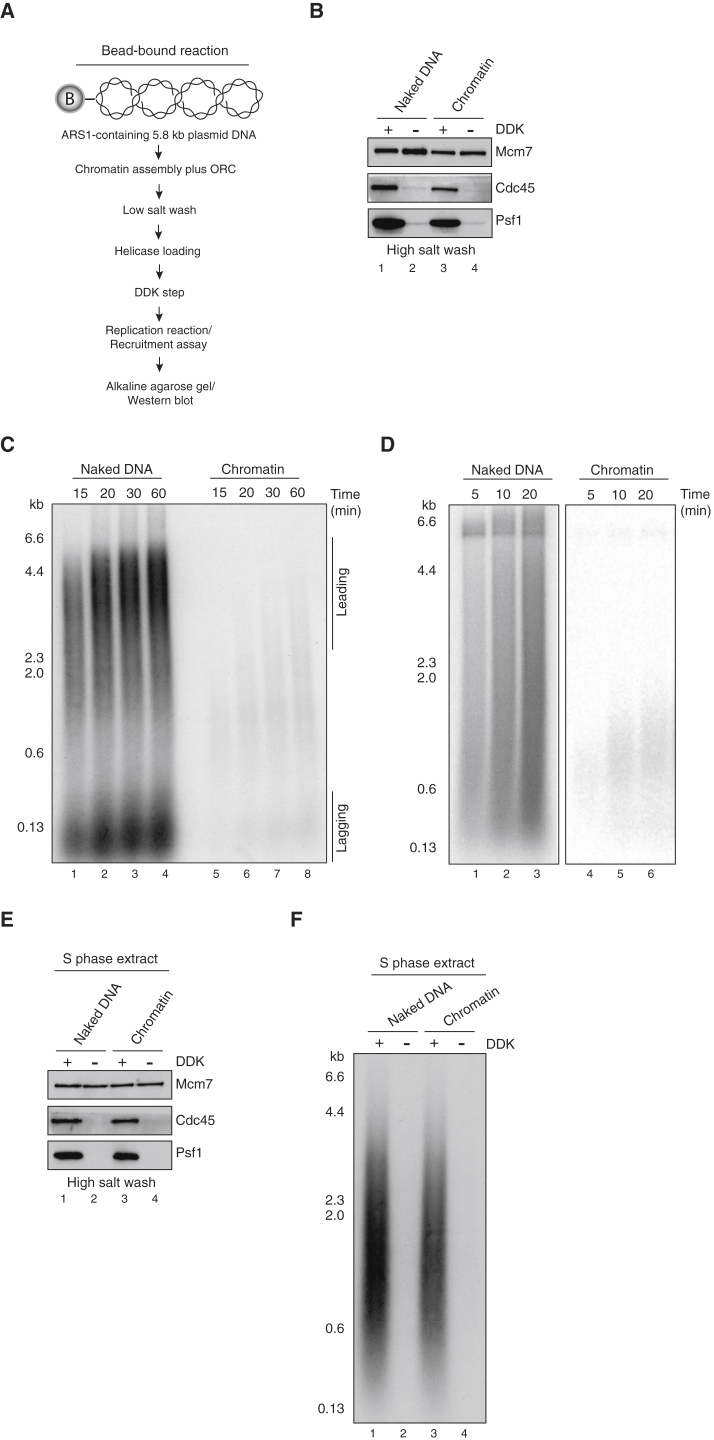
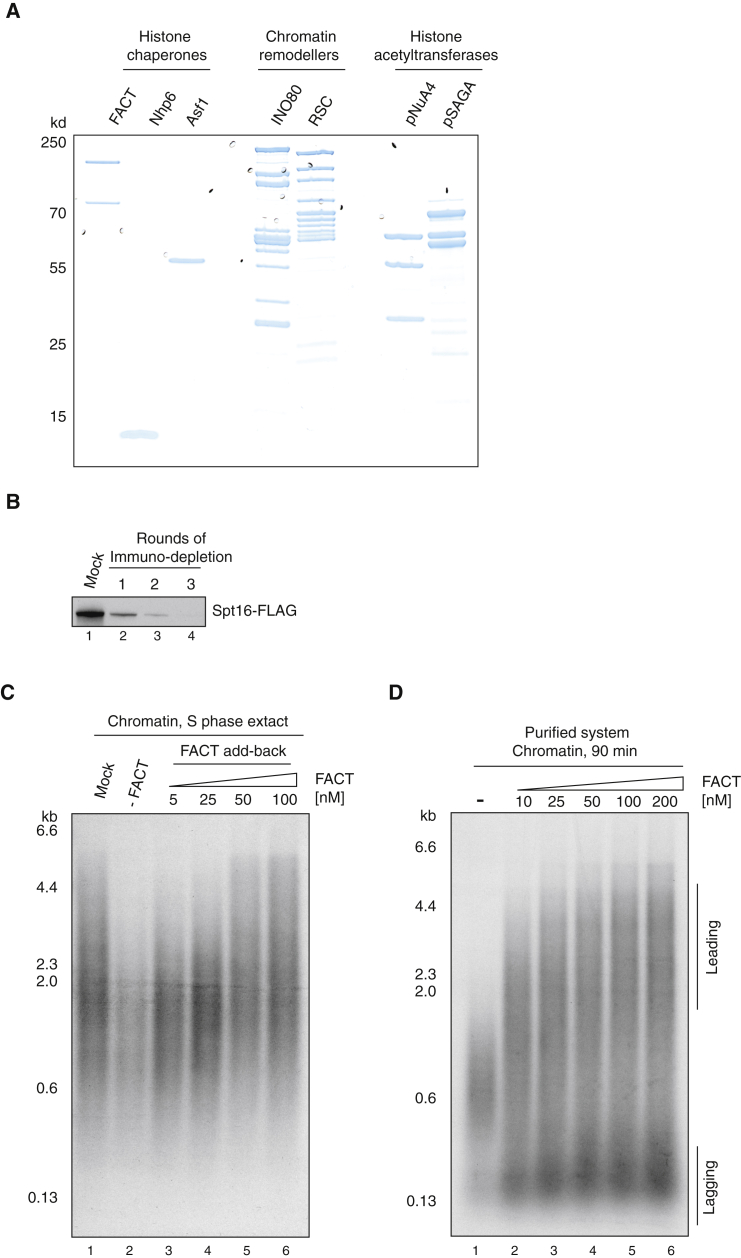
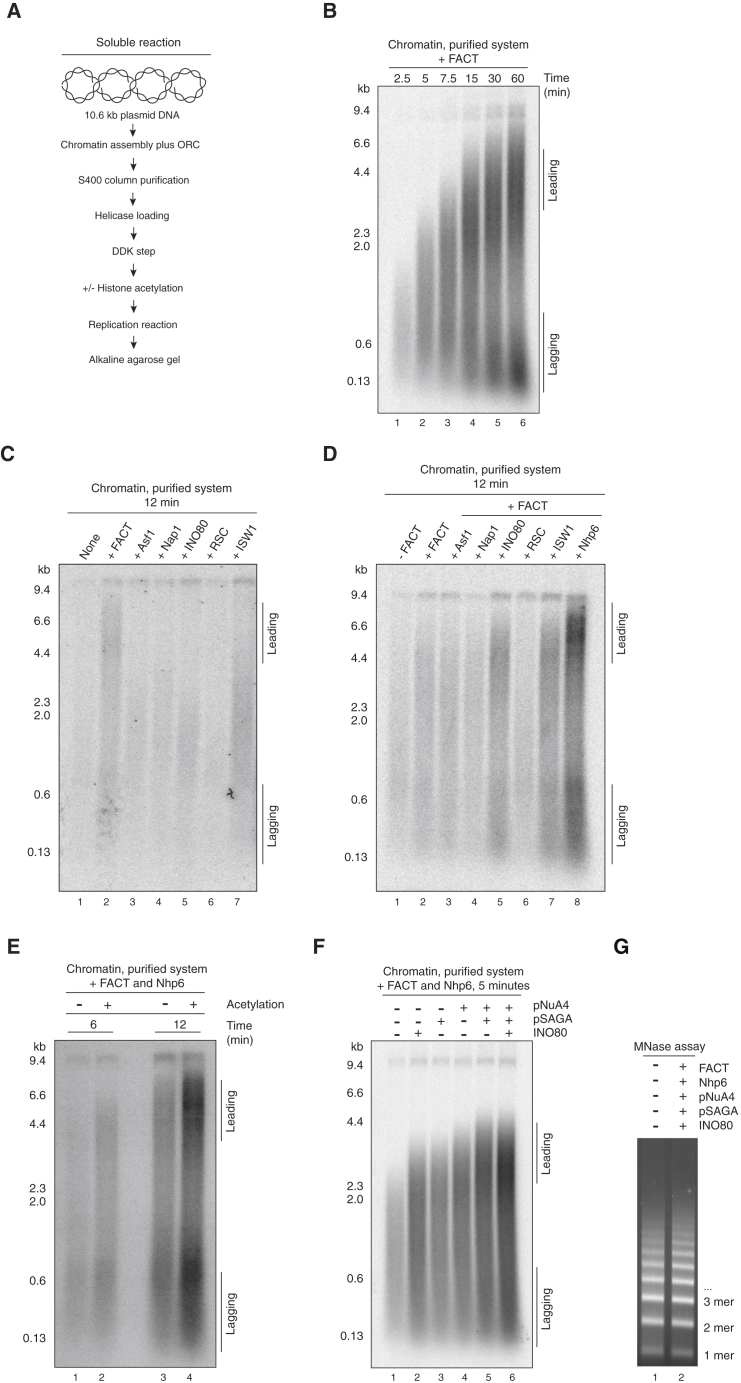
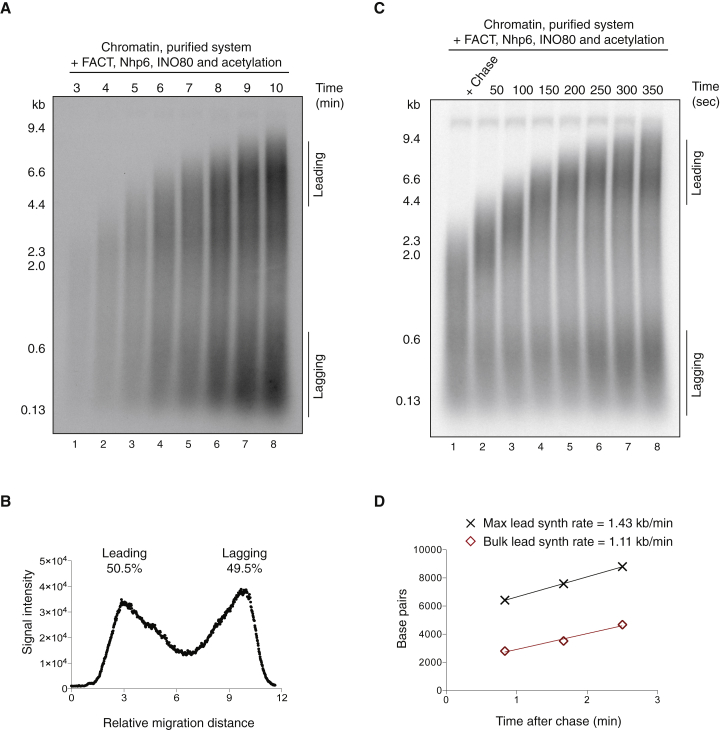
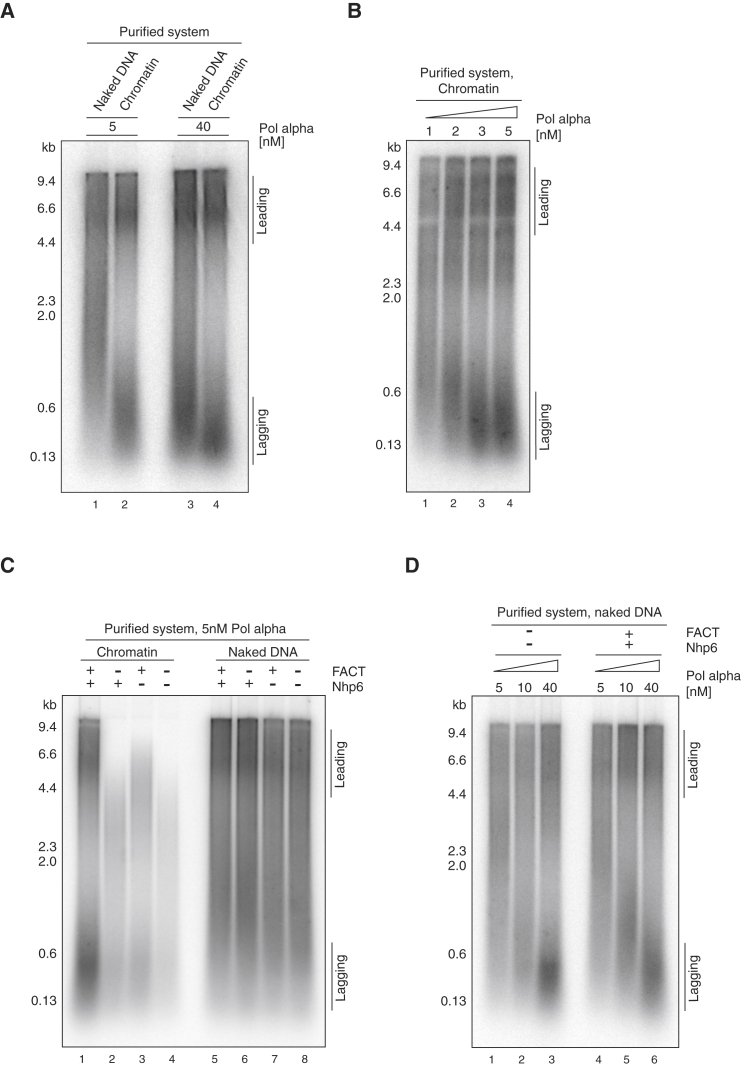
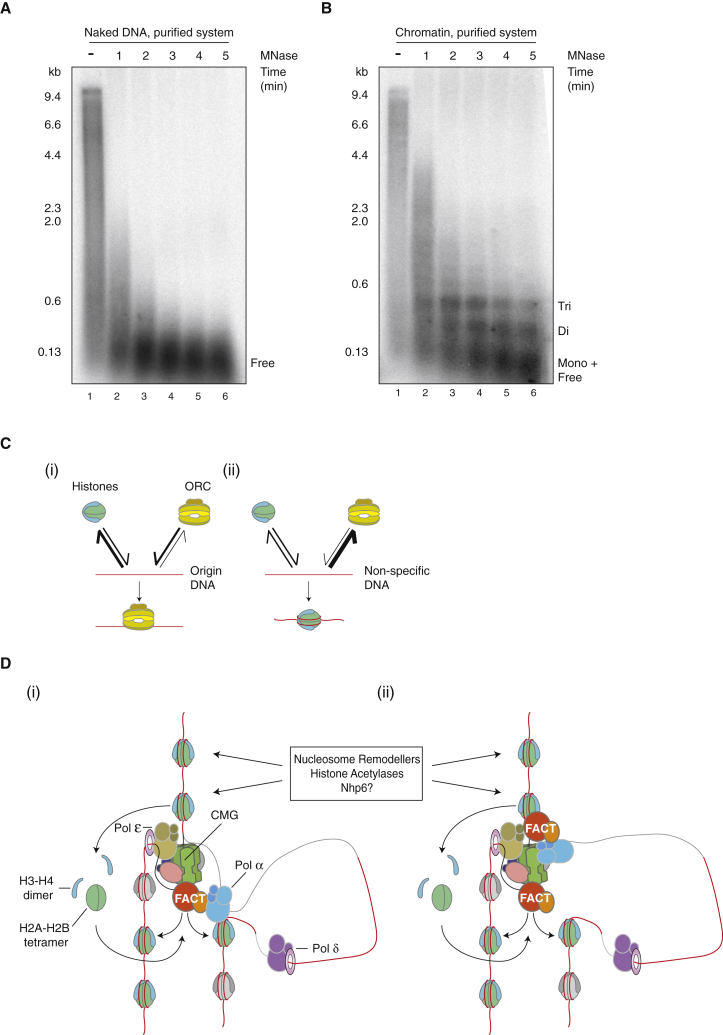
The integrity of eukaryotic genomes requires rapid and regulated chromatin replication. How this is accomplished is still poorly understood. Using purified yeast replication proteins and fully chromatinized templates, we have reconstituted this process in vitro. We show that chromatin enforces DNA replication origin specificity by preventing non-specific MCM helicase loading. Helicase activation occurs efficiently in the context of chromatin, but subsequent replisome progression requires the histone chaperone FACT (facilitates chromatin transcription). The FACT-associated Nhp6 protein, the nucleosome remodelers INO80 or ISW1A, and the lysine acetyltransferases Gcn5 and Esa1 each contribute separately to maximum DNA synthesis rates. Chromatin promotes the regular priming of lagging-strand DNA synthesis by facilitating DNA polymerase α function at replication forks. Finally, nucleosomes disrupted during replication are efficiently re-assembled into regular arrays on nascent DNA. Our work defines the minimum requirements for chromatin replication in vitro and shows how multiple chromatin factors might modulate replication fork rates in vivo.
Keywords: DNA replication; biochemistry; chromatin.
Copyright © 2017 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
