

M1 muscarinic allosteric modulators slow prion neurodegeneration and restore memory loss
- PMID: 27991860
- PMCID: PMC5272187
- DOI: 10.1172/JCI87526
M1 muscarinic allosteric modulators slow prion neurodegeneration and restore memory loss
Abstract
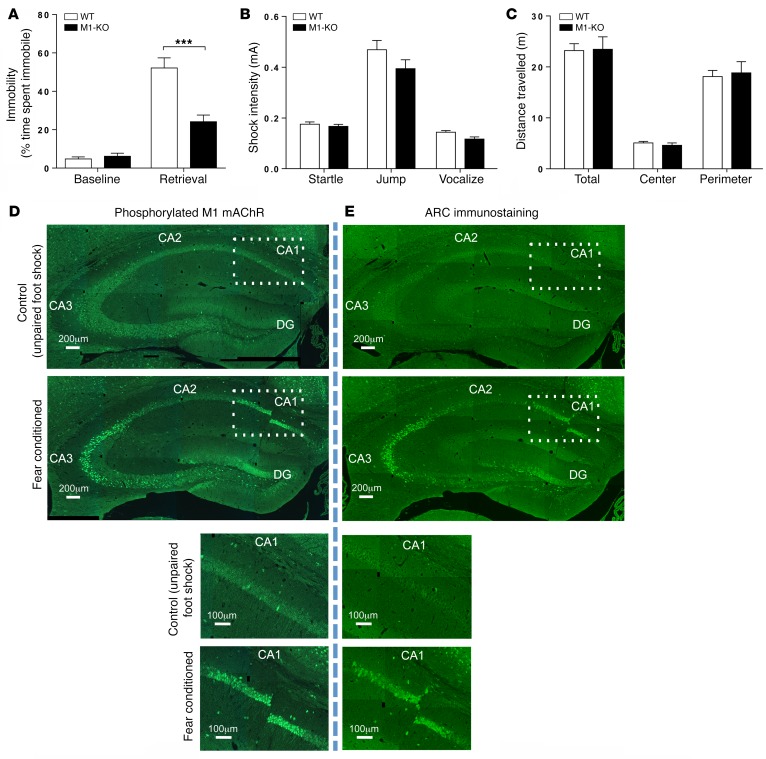
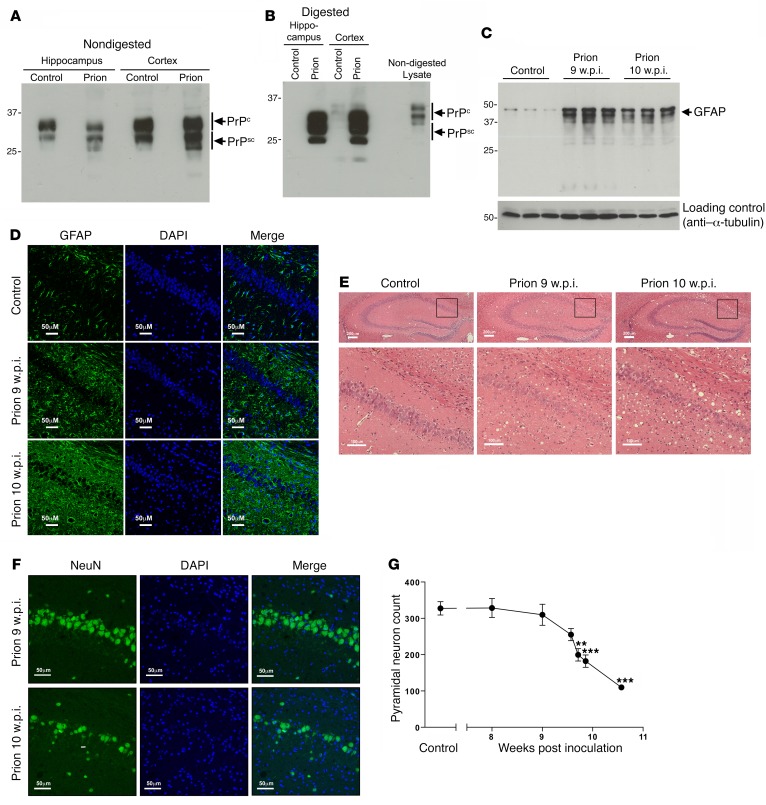
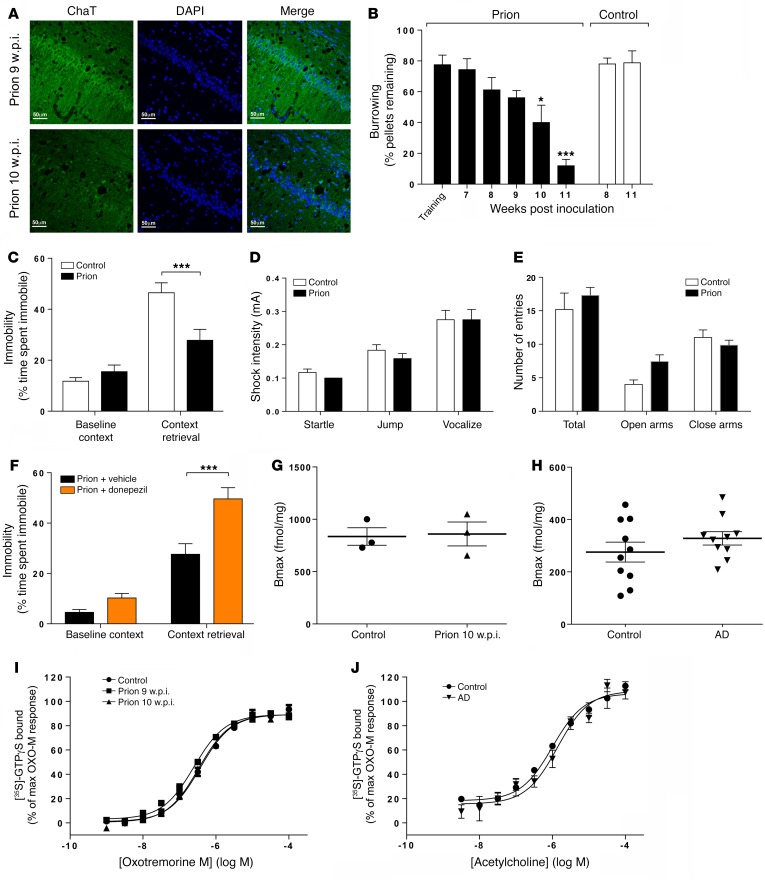
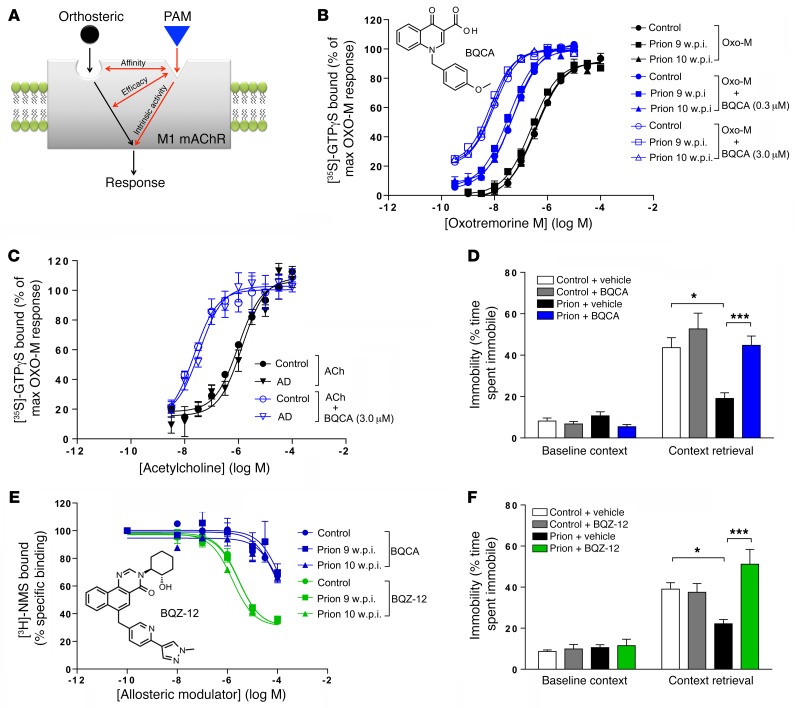
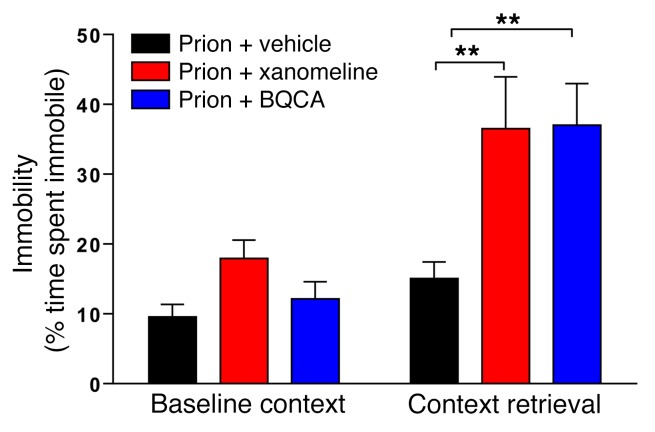
The current frontline symptomatic treatment for Alzheimer's disease (AD) is whole-body upregulation of cholinergic transmission via inhibition of acetylcholinesterase. This approach leads to profound dose-related adverse effects. An alternative strategy is to selectively target muscarinic acetylcholine receptors, particularly the M1 muscarinic acetylcholine receptor (M1 mAChR), which was previously shown to have procognitive activity. However, developing M1 mAChR-selective orthosteric ligands has proven challenging. Here, we have shown that mouse prion disease shows many of the hallmarks of human AD, including progressive terminal neurodegeneration and memory deficits due to a disruption of hippocampal cholinergic innervation. The fact that we also show that muscarinic signaling is maintained in both AD and mouse prion disease points to the latter as an excellent model for testing the efficacy of muscarinic pharmacological entities. The memory deficits we observed in mouse prion disease were completely restored by treatment with benzyl quinolone carboxylic acid (BQCA) and benzoquinazoline-12 (BQZ-12), two highly selective positive allosteric modulators (PAMs) of M1 mAChRs. Furthermore, prolonged exposure to BQCA markedly extended the lifespan of diseased mice. Thus, enhancing hippocampal muscarinic signaling using M1 mAChR PAMs restored memory loss and slowed the progression of mouse prion disease, indicating that this ligand type may have clinical benefit in diseases showing defective cholinergic transmission, such as AD.
Conflict of interest statement
The authors have declared that no conflict of interest exists.
Figures


References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
