

Arabidopsis SWI/SNF chromatin remodeling complex binds both promoters and terminators to regulate gene expression
- PMID: 27994035
- PMCID: PMC5389626
- DOI: 10.1093/nar/gkw1273
Arabidopsis SWI/SNF chromatin remodeling complex binds both promoters and terminators to regulate gene expression
Abstract
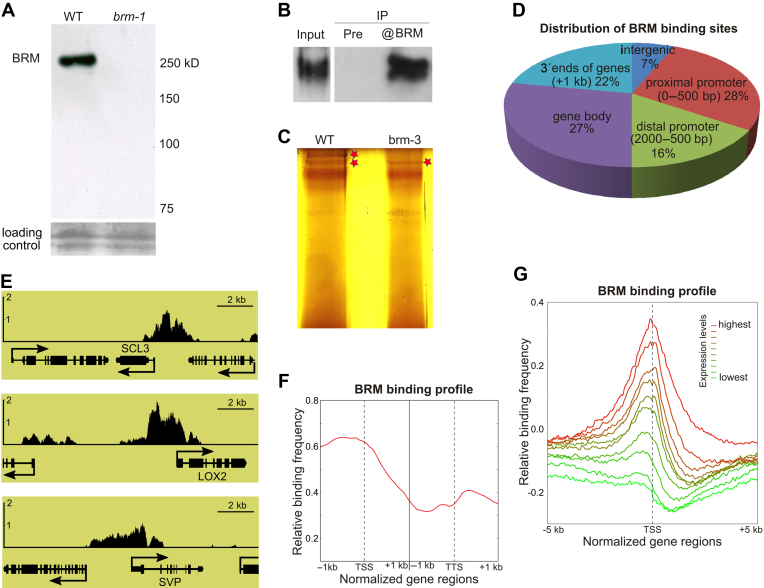
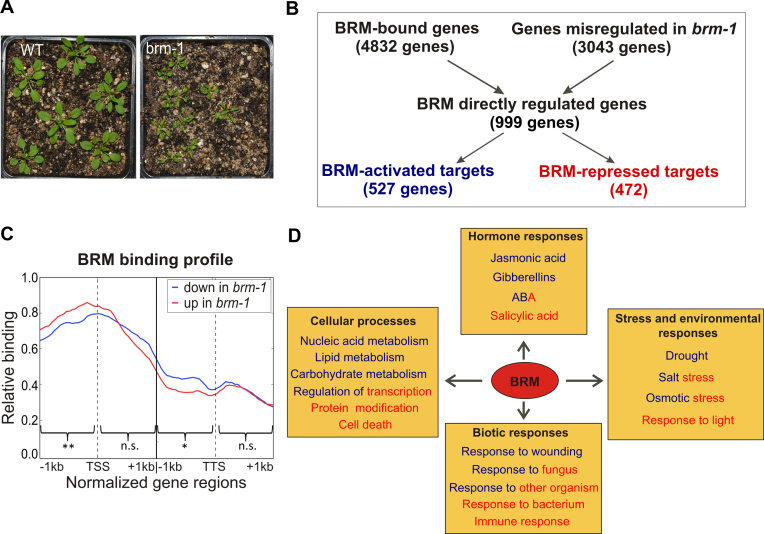
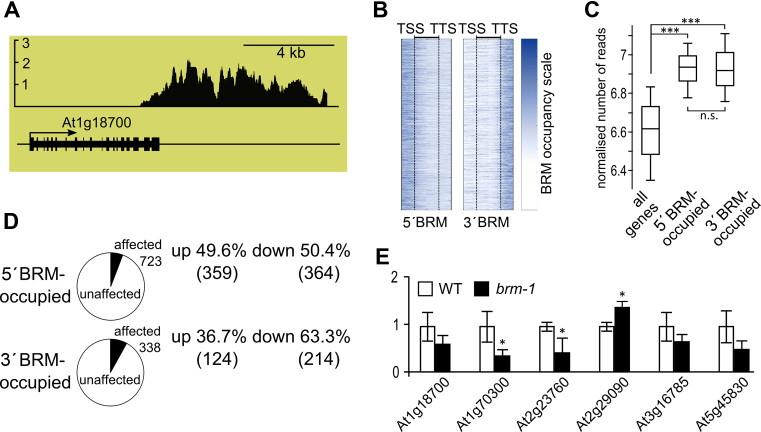
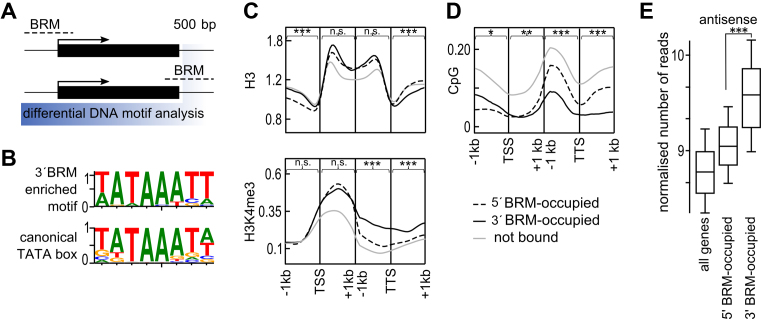
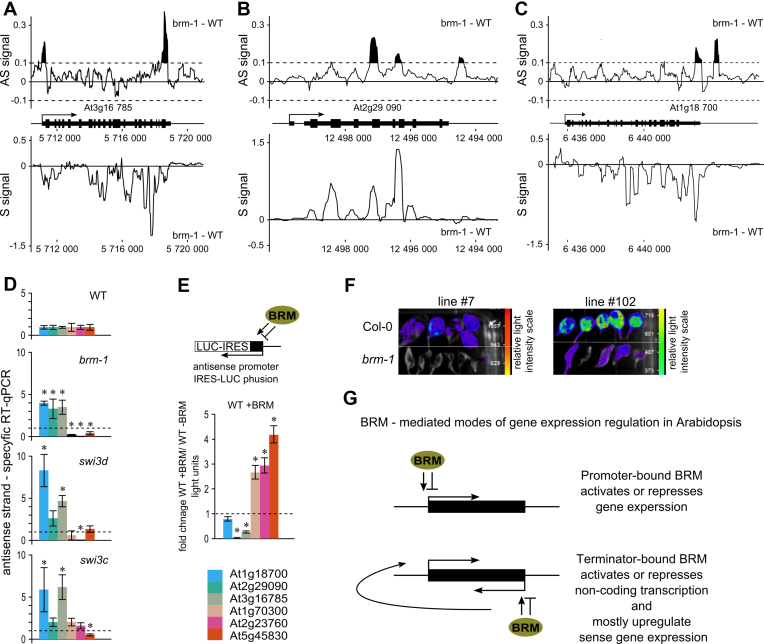
ATP-dependent chromatin remodeling complexes are important regulators of gene expression in Eukaryotes. In plants, SWI/SNF-type complexes have been shown critical for transcriptional control of key developmental processes, growth and stress responses. To gain insight into mechanisms underlying these roles, we performed whole genome mapping of the SWI/SNF catalytic subunit BRM in Arabidopsis thaliana, combined with transcript profiling experiments. Our data show that BRM occupies thousands of sites in Arabidopsis genome, most of which located within or close to genes. Among identified direct BRM transcriptional targets almost equal numbers were up- and downregulated upon BRM depletion, suggesting that BRM can act as both activator and repressor of gene expression. Interestingly, in addition to genes showing canonical pattern of BRM enrichment near transcription start site, many other genes showed a transcription termination site-centred BRM occupancy profile. We found that BRM-bound 3΄ gene regions have promoter-like features, including presence of TATA boxes and high H3K4me3 levels, and possess high antisense transcriptional activity which is subjected to both activation and repression by SWI/SNF complex. Our data suggest that binding to gene terminators and controlling transcription of non-coding RNAs is another way through which SWI/SNF complex regulates expression of its targets.
© The Author(s) 2016. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
