

Modeling Protein Excited-state Structures from "Over-length" Chemical Cross-links
- PMID: 27994050
- PMCID: PMC5270465
- DOI: 10.1074/jbc.M116.761841
Modeling Protein Excited-state Structures from "Over-length" Chemical Cross-links
Abstract
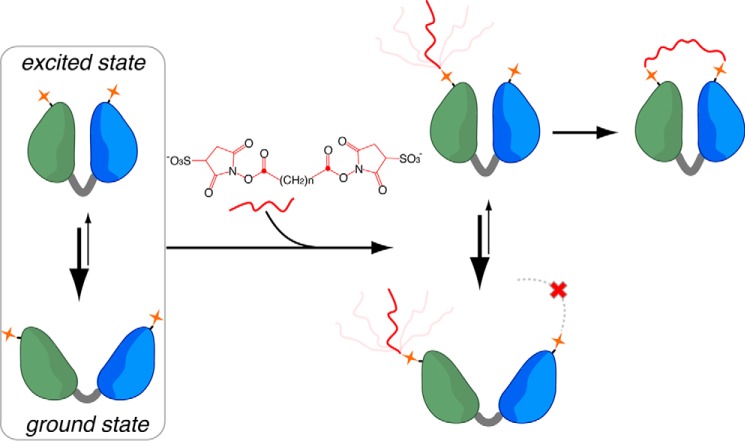
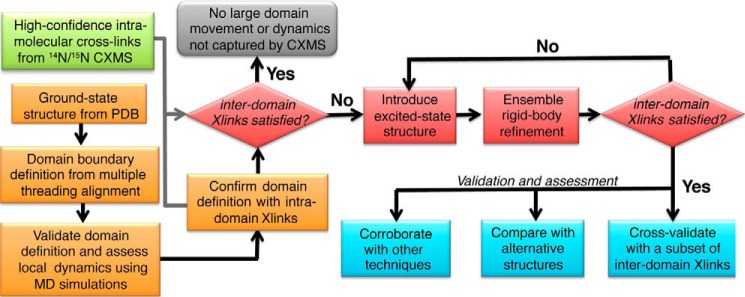
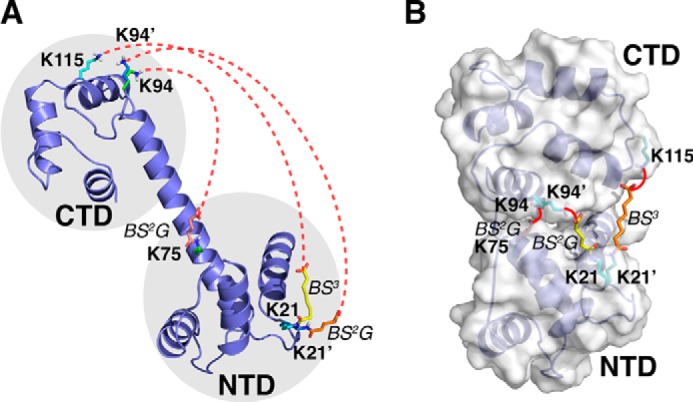
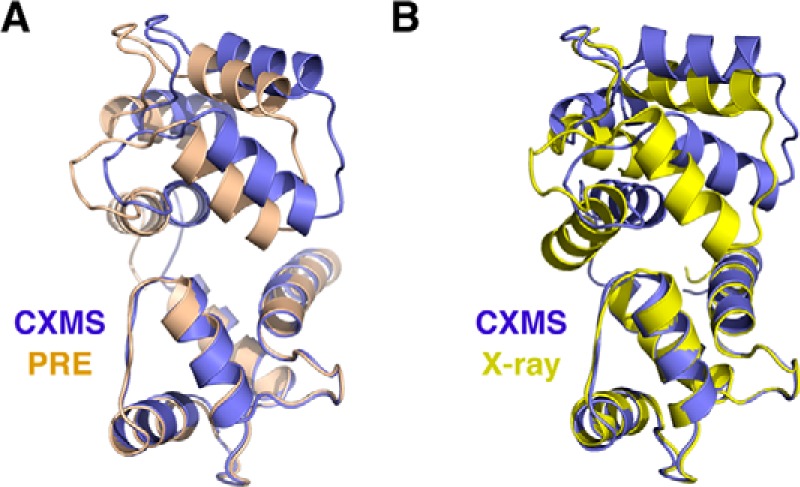
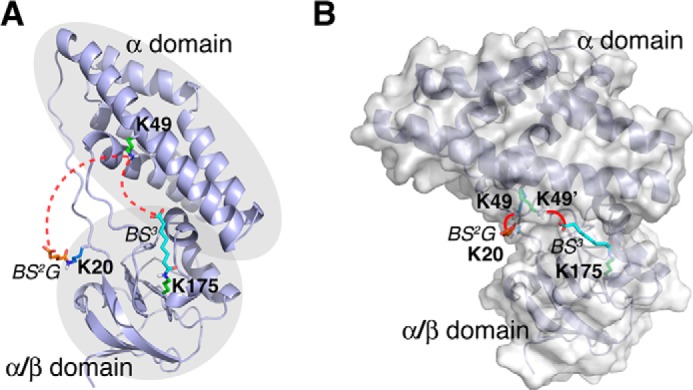
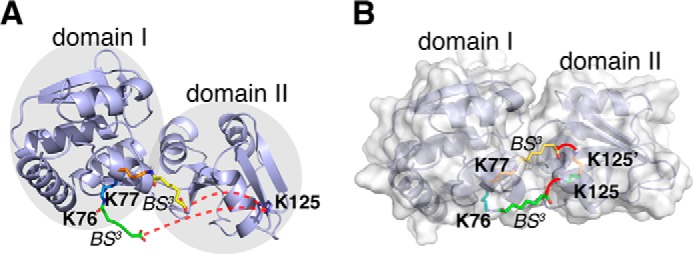
Chemical cross-linking coupled with mass spectroscopy (CXMS) provides proximity information for the cross-linked residues and is used increasingly for modeling protein structures. However, experimentally identified cross-links are sometimes incompatible with the known structure of a protein, as the distance calculated between the cross-linked residues far exceeds the maximum length of the cross-linker. The discrepancies may persist even after eliminating potentially false cross-links and excluding intermolecular ones. Thus the "over-length" cross-links may arise from alternative excited-state conformation of the protein. Here we present a method and associated software DynaXL for visualizing the ensemble structures of multidomain proteins based on intramolecular cross-links identified by mass spectrometry with high confidence. Representing the cross-linkers and cross-linking reactions explicitly, we show that the protein excited-state structure can be modeled with as few as two over-length cross-links. We demonstrate the generality of our method with three systems: calmodulin, enzyme I, and glutamine-binding protein, and we show that these proteins alternate between different conformations for interacting with other proteins and ligands. Taken together, the over-length chemical cross-links contain valuable information about protein dynamics, and our findings here illustrate the relationship between dynamic domain movement and protein function.
Keywords: calmodulin (CaM); mass spectrometry (MS); molecular dynamics; protein cross-linking; protein domain; protein dynamics; protein excited state; structural model.
© 2017 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
