

Disrupting the blood-brain barrier by focused ultrasound induces sterile inflammation
- PMID: 27994152
- PMCID: PMC5224365
- DOI: 10.1073/pnas.1614777114
Disrupting the blood-brain barrier by focused ultrasound induces sterile inflammation
Abstract
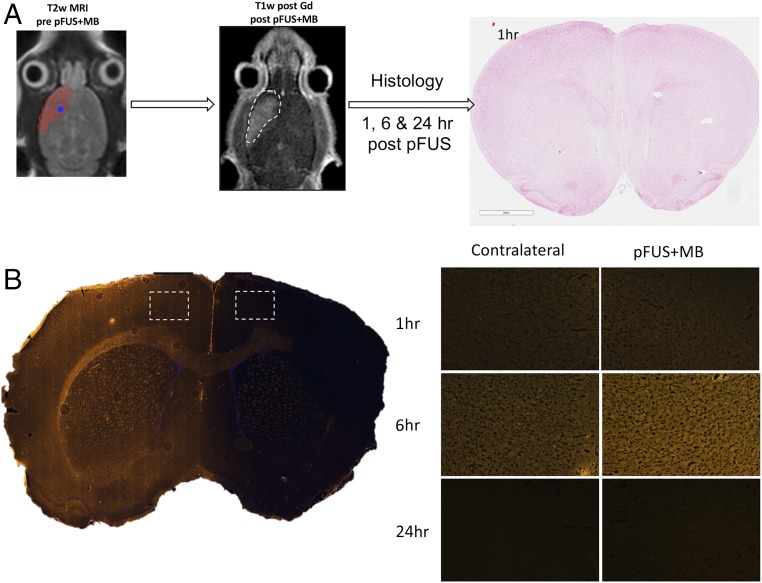
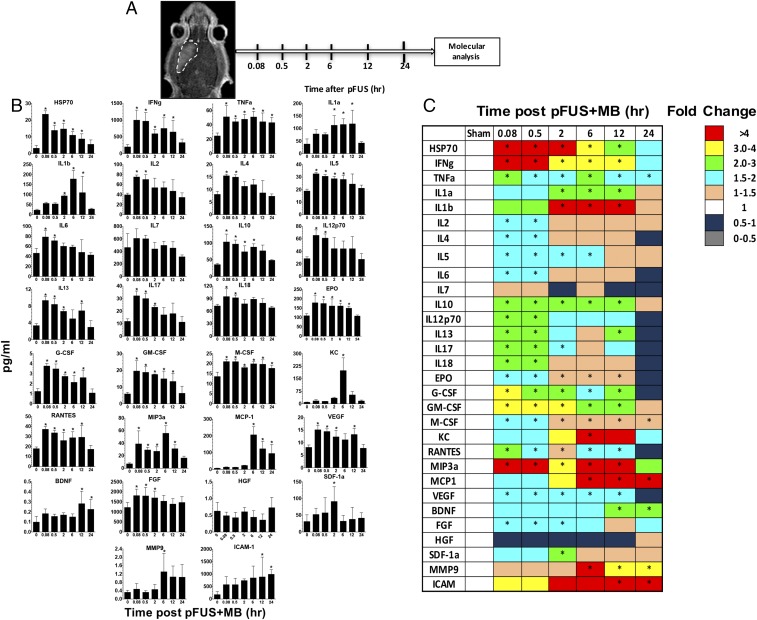
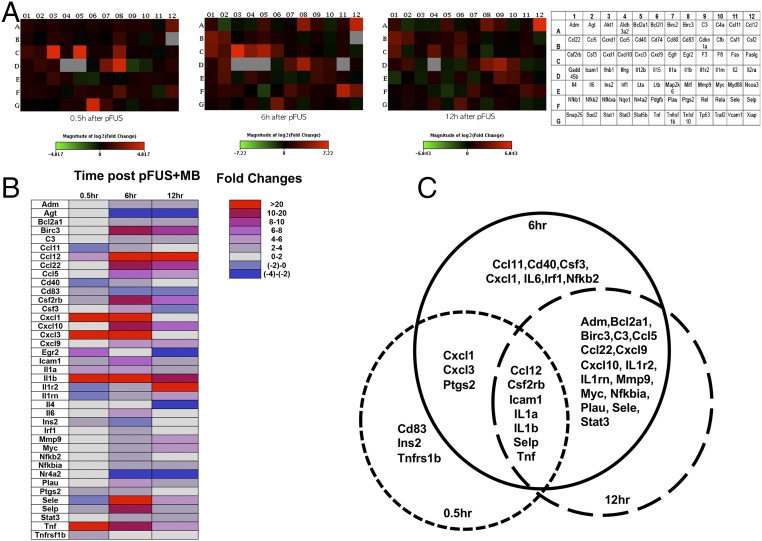
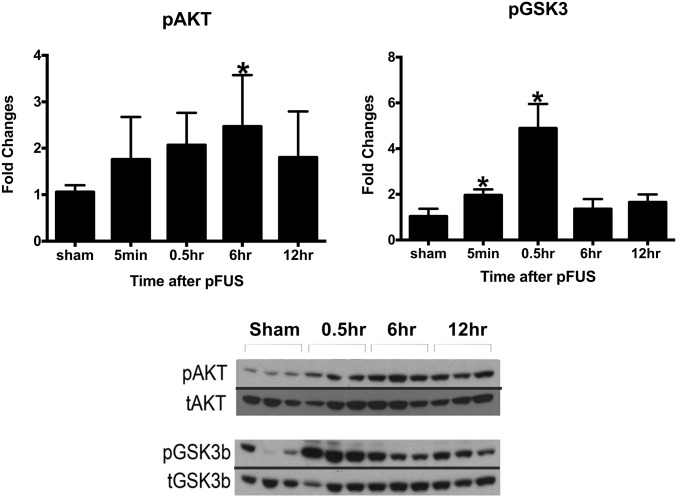
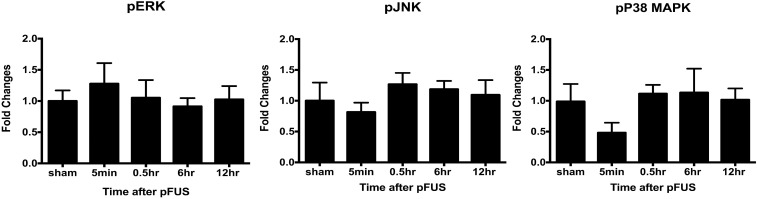
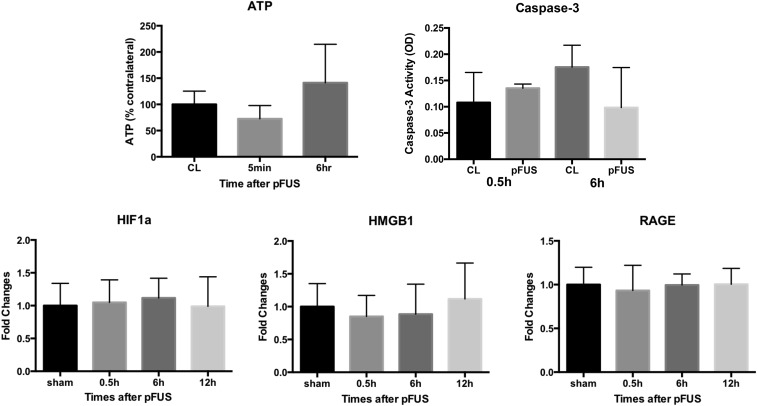
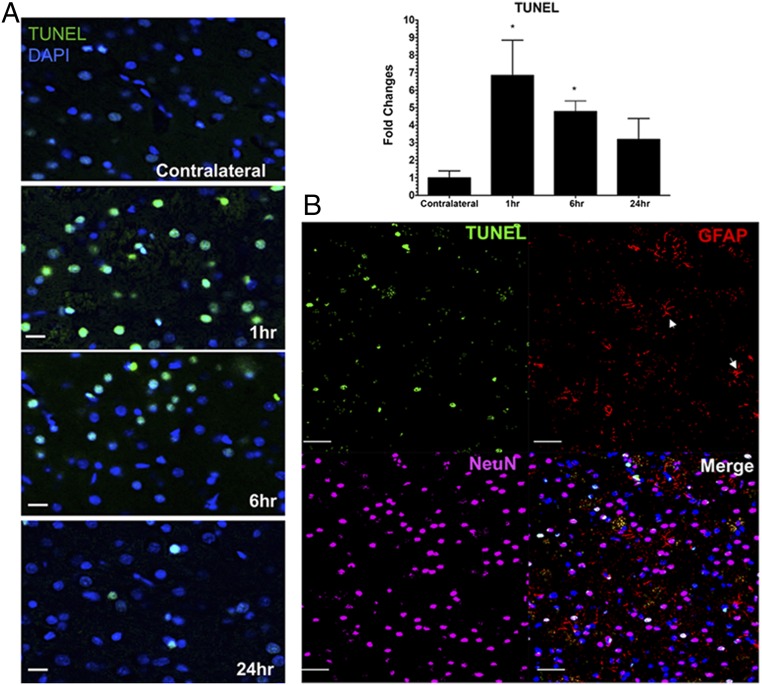
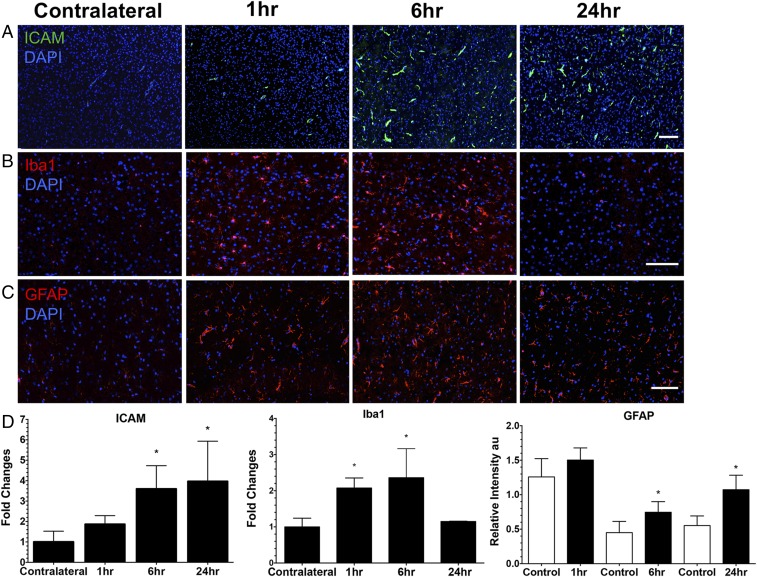
MRI-guided pulsed focused ultrasound (pFUS) combined with systemic infusion of ultrasound contrast agent microbubbles (MB) causes localized blood-brain barrier (BBB) disruption that is currently being advocated for increasing drug or gene delivery in neurological diseases. The mechanical acoustic cavitation effects of opening the BBB by low-intensity pFUS+MB, as evidenced by contrast-enhanced MRI, resulted in an immediate damage-associated molecular pattern (DAMP) response including elevations in heat-shock protein 70, IL-1, IL-18, and TNFα indicative of a sterile inflammatory response (SIR) in the parenchyma. Concurrent with DAMP presentation, significant elevations in proinflammatory, antiinflammatory, and trophic factors along with neurotrophic and neurogenesis factors were detected; these elevations lasted 24 h. Transcriptomic analysis of sonicated brain supported the proteomic findings and indicated that the SIR was facilitated through the induction of the NFκB pathway. Histological evaluation demonstrated increased albumin in the parenchyma that cleared by 24 h along with TUNEL+ neurons, activated astrocytes, microglia, and increased cell adhesion molecules in the vasculature. Infusion of fluorescent beads 3 d before pFUS+MB revealed the infiltration of CD68+ macrophages at 6 d postsonication, as is consistent with an innate immune response. pFUS+MB is being considered as part of a noninvasive adjuvant treatment for malignancy or neurodegenerative diseases. These results demonstrate that pFUS+MB induces an SIR compatible with ischemia or mild traumatic brain injury. Further investigation will be required before this approach can be widely implemented in clinical trials.
Keywords: blood-brain barrier; magnetic resonance imaging; microbubbles; pulsed focused ultrasound; sterile inflammation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
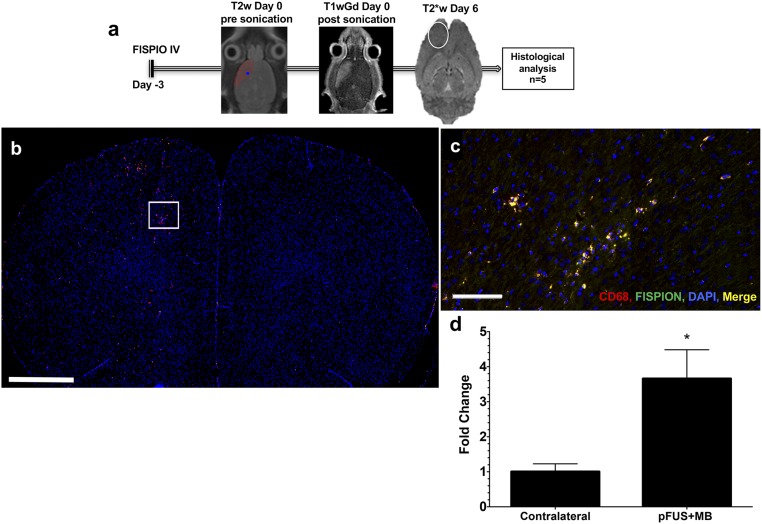
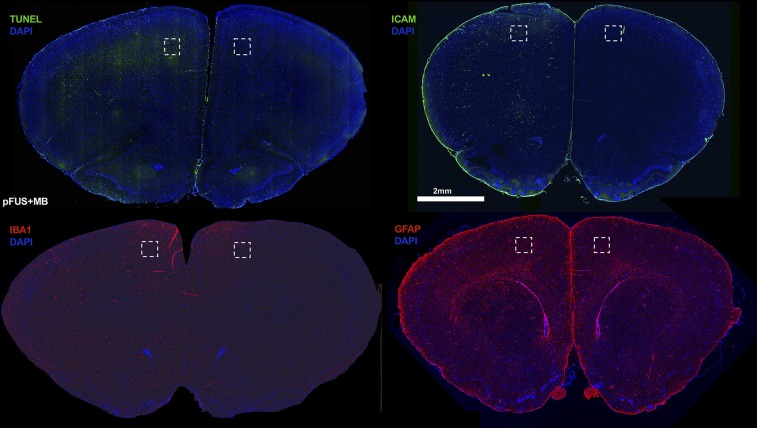
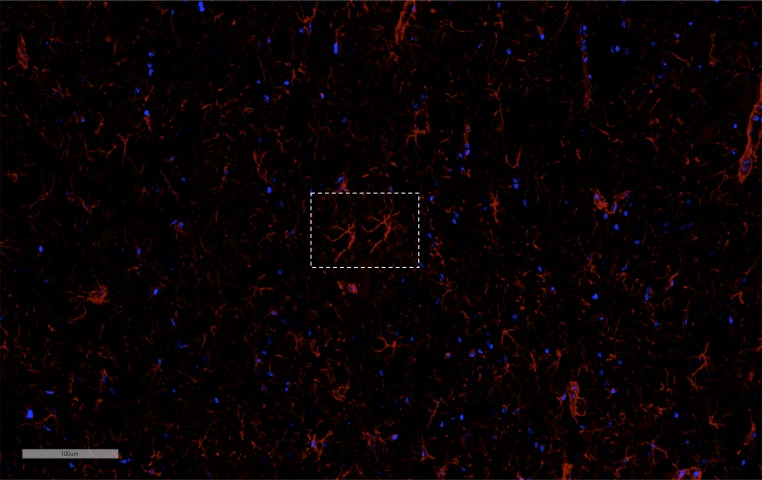
Comment in
-
A sterile animal model for neuroinflammation?Sci Transl Med. 2017 Jan 18;9(373):eaal4994. doi: 10.1126/scitranslmed.aal4994. Sci Transl Med. 2017. PMID: 28100836
References
-
- Schoknecht K, David Y, Heinemann U. The blood-brain barrier-gatekeeper to neuronal homeostasis: Clinical implications in the setting of stroke. Semin Cell Dev Biol. 2015;38:35–42. - PubMed
-
- Cernak I, O’Connor C, Vink R. Inhibition of cyclooxygenase 2 by nimesulide improves cognitive outcome more than motor outcome following diffuse traumatic brain injury in rats. Exp Brain Res. 2002;147(2):193–199. - PubMed
-
- Denes A, Thornton P, Rothwell NJ, Allan SM. Inflammation and brain injury: Acute cerebral ischaemia, peripheral and central inflammation. Brain Behav Immun. 2010;24(5):708–723. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
