

Identification of miR-31-5p, miR-141-3p, miR-200c-3p, and GLT1 as human liver aging markers sensitive to donor-recipient age-mismatch in transplants
- PMID: 27995756
- PMCID: PMC5334540
- DOI: 10.1111/acel.12549
Identification of miR-31-5p, miR-141-3p, miR-200c-3p, and GLT1 as human liver aging markers sensitive to donor-recipient age-mismatch in transplants
Abstract
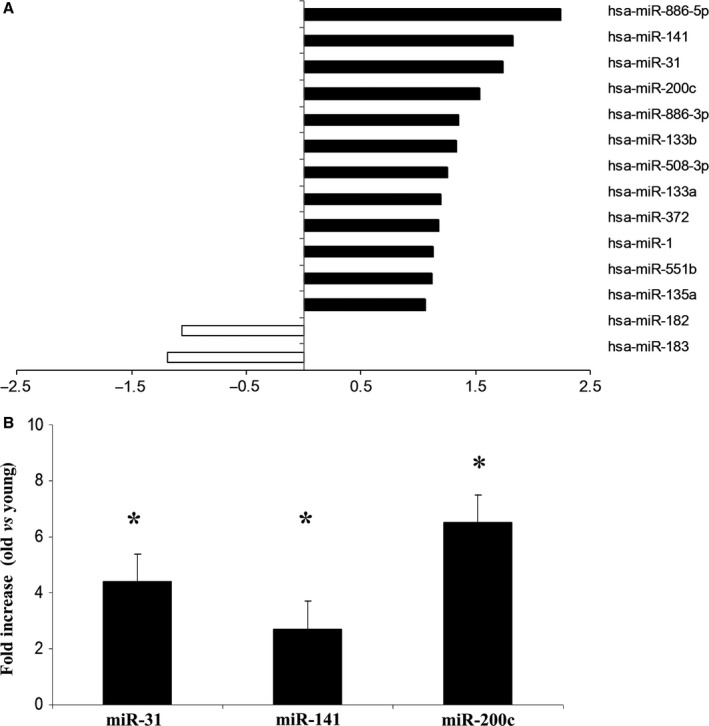
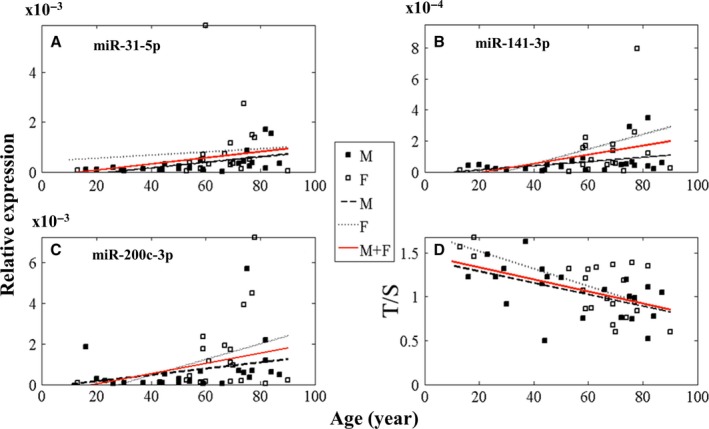
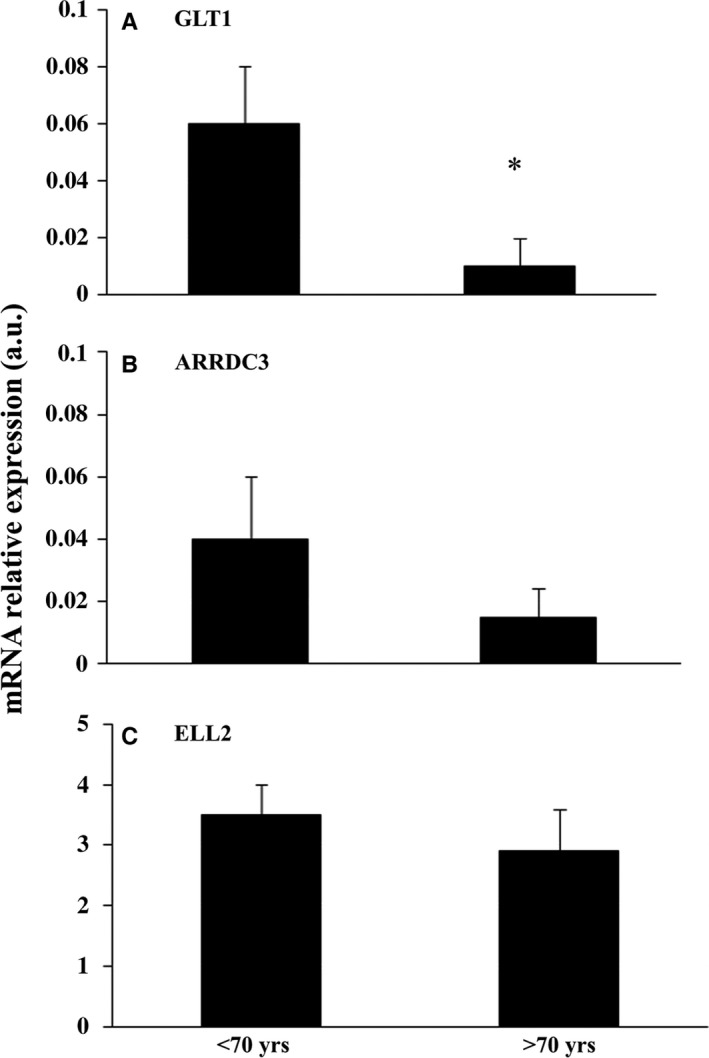
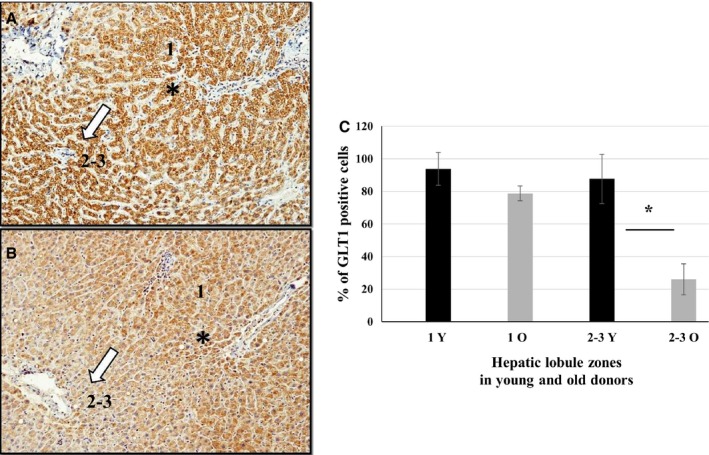
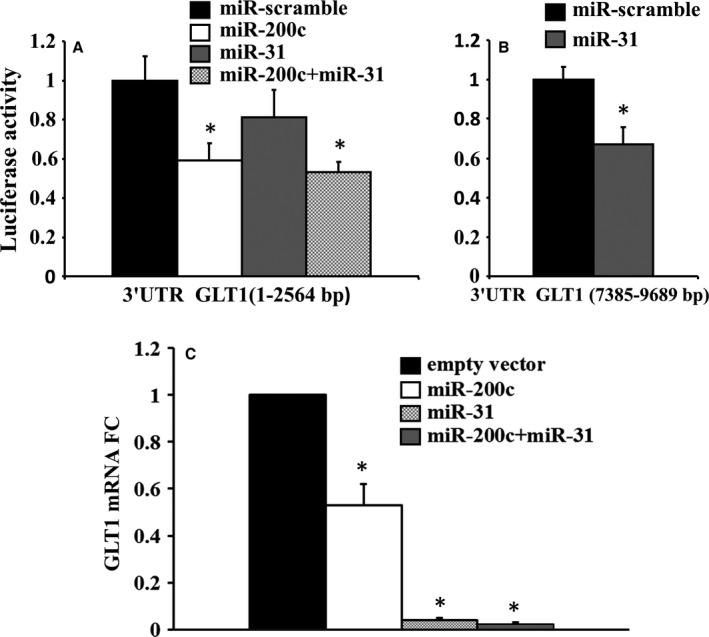
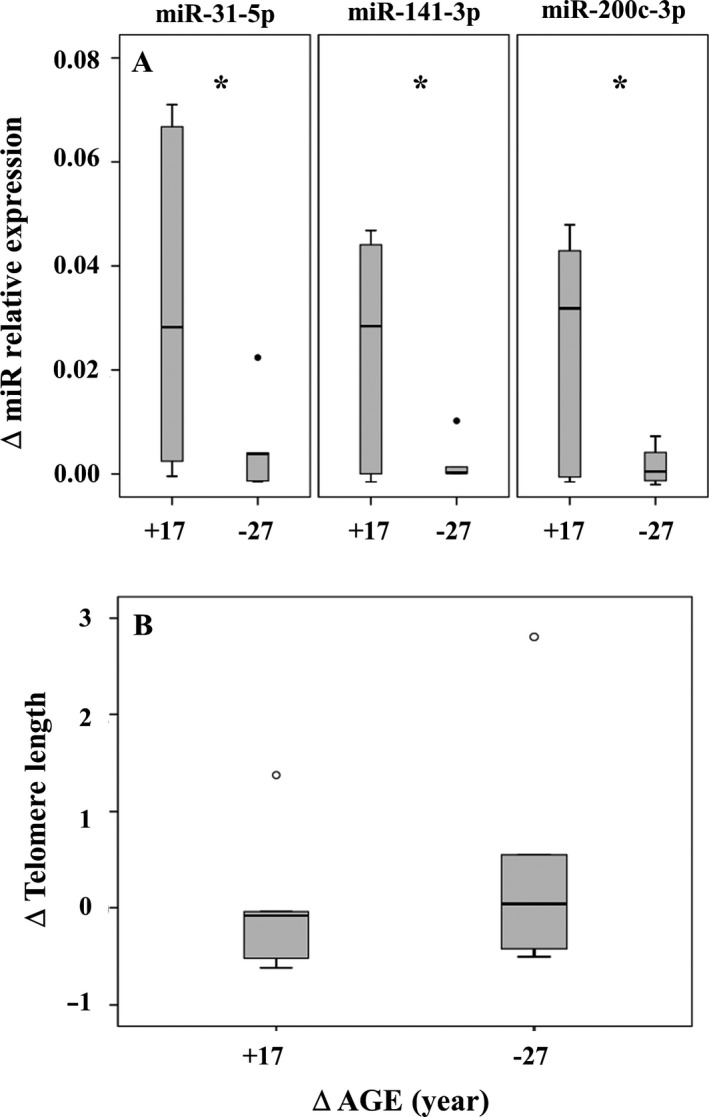
To understand why livers from aged donors are successfully used for transplants, we looked for markers of liver aging in 71 biopsies from donors aged 12-92 years before transplants and in 11 biopsies after transplants with high donor-recipient age-mismatch. We also assessed liver function in 36 age-mismatched recipients. The major findings were the following: (i) miR-31-5p, miR-141-3p, and miR-200c-3p increased with age, as assessed by microRNAs (miRs) and mRNA transcript profiling in 12 biopsies and results were validated by RT-qPCR in a total of 58 biopsies; (ii) telomere length measured by qPCR in 45 samples showed a significant age-dependent shortage; (iii) a bioinformatic approach combining transcriptome and miRs data identified putative miRs targets, the most informative being GLT1, a glutamate transporter expressed in hepatocytes. GLT1 was demonstrated by luciferase assay to be a target of miR-31-5p and miR-200c-3p, and both its mRNA (RT-qPCR) and protein (immunohistochemistry) significantly decreased with age in liver biopsies and in hepatic centrilobular zone, respectively; (iv) miR-31-5p, miR-141-3p and miR-200c-3p expression was significantly affected by recipient age (older environment) as assessed in eleven cases of donor-recipient extreme age-mismatch; (v) the analysis of recipients plasma by N-glycans profiling, capable of assessing liver functions and biological age, showed that liver function recovered after transplants, independently of age-mismatch, and recipients apparently 'rejuvenated' according to their glycomic age. In conclusion, we identified new markers of aging in human liver, their relevance in donor-recipient age-mismatches in transplantation, and offered positive evidence for the use of organs from old donors.
Keywords: GLT1; N-glycans; age-mismatches; allograft; elderly donors; microRNAs; telomere length.
© 2016 The Authors. Aging Cell published by the Anatomical Society and John Wiley & Sons Ltd.
Figures

References
-
- Albertini MC, Olivieri F, Lazzarini R, Pilolli F, Galli F, Spada G, Accorsi A, Rippo MR, Procopio AD (2011) Predicting microRNA modulation in human prostate cancer using a simple String IDentifier (SID1.0). J. Biomed. Inform. 44, 615–620. - PubMed
-
- Banaudha K, Kaliszewski M, Korolnek T, Florea L, Yeung ML, Jeang KT, Kumar A (2011) MicroRNA silencing of tumor suppressor DLC‐1 promotes efficient hepatitis C virus replication in primary human hepatocytes. Hepatology 53, 53–61. - PubMed
-
- Berger UV, Hediger MA (2006) Distribution of the glutamate transporters GLT‐1 (SLC1A2) and GLAST (SLC1A3) in peripheral organs. Anat. Embryol. 211, 595–606. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
