

Central Role of CD169+ Lymph Node Resident Macrophages in the Adjuvanticity of the QS-21 Component of AS01
- PMID: 27996000
- PMCID: PMC5172233
- DOI: 10.1038/srep39475
Central Role of CD169+ Lymph Node Resident Macrophages in the Adjuvanticity of the QS-21 Component of AS01
Abstract
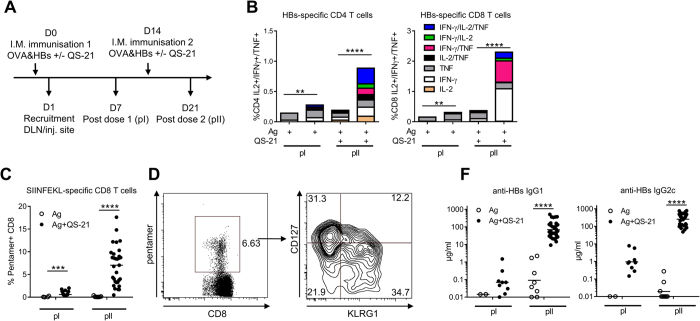
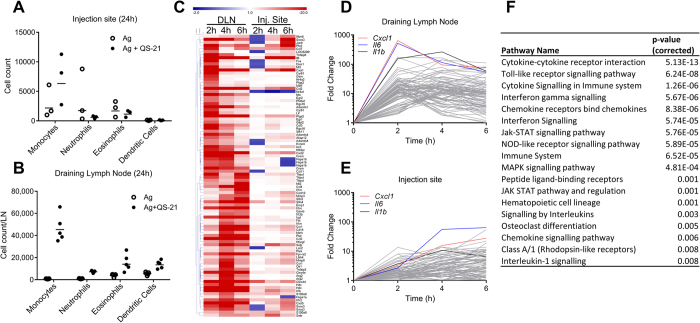
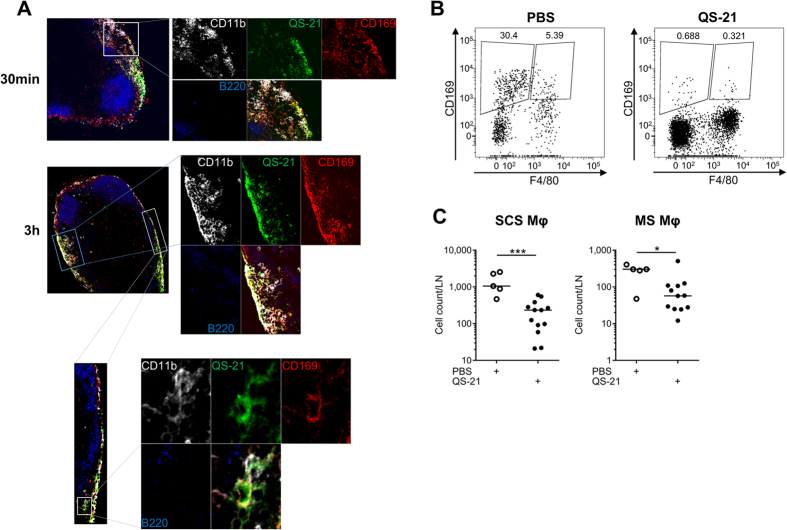
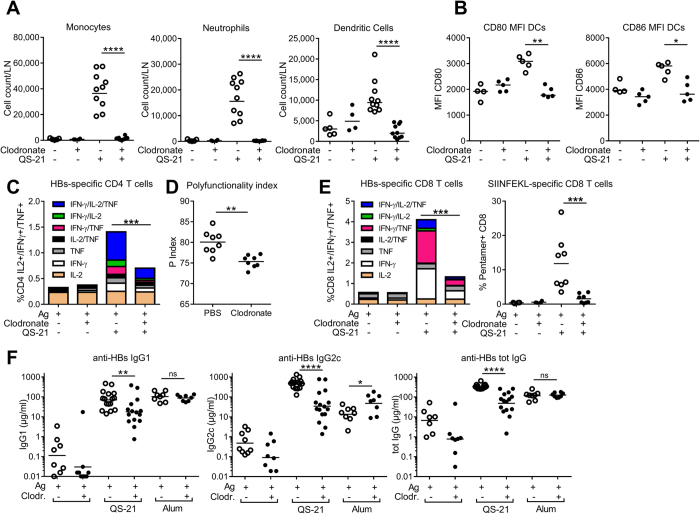
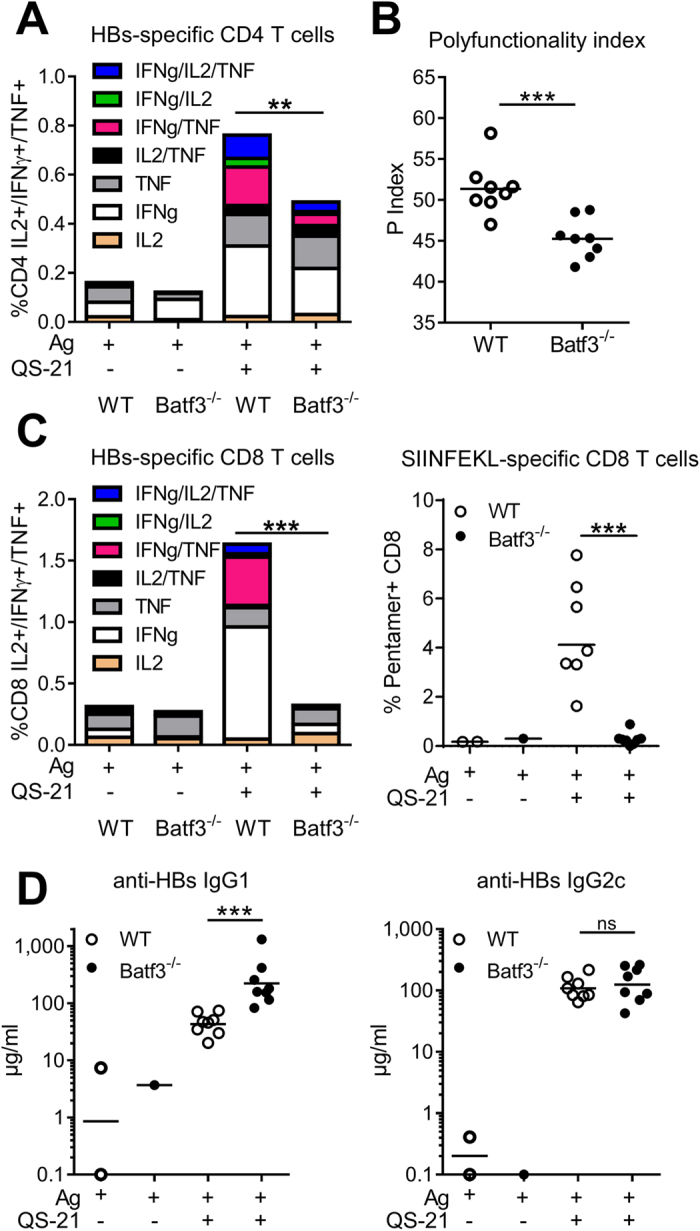
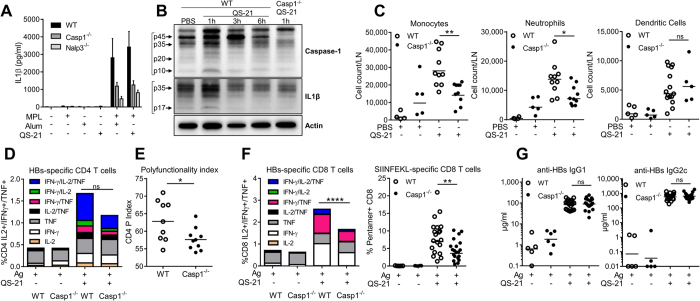
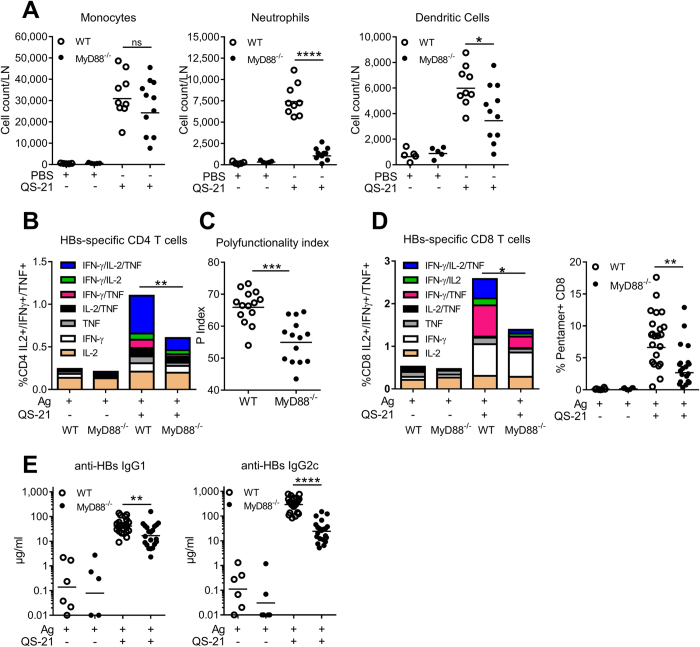
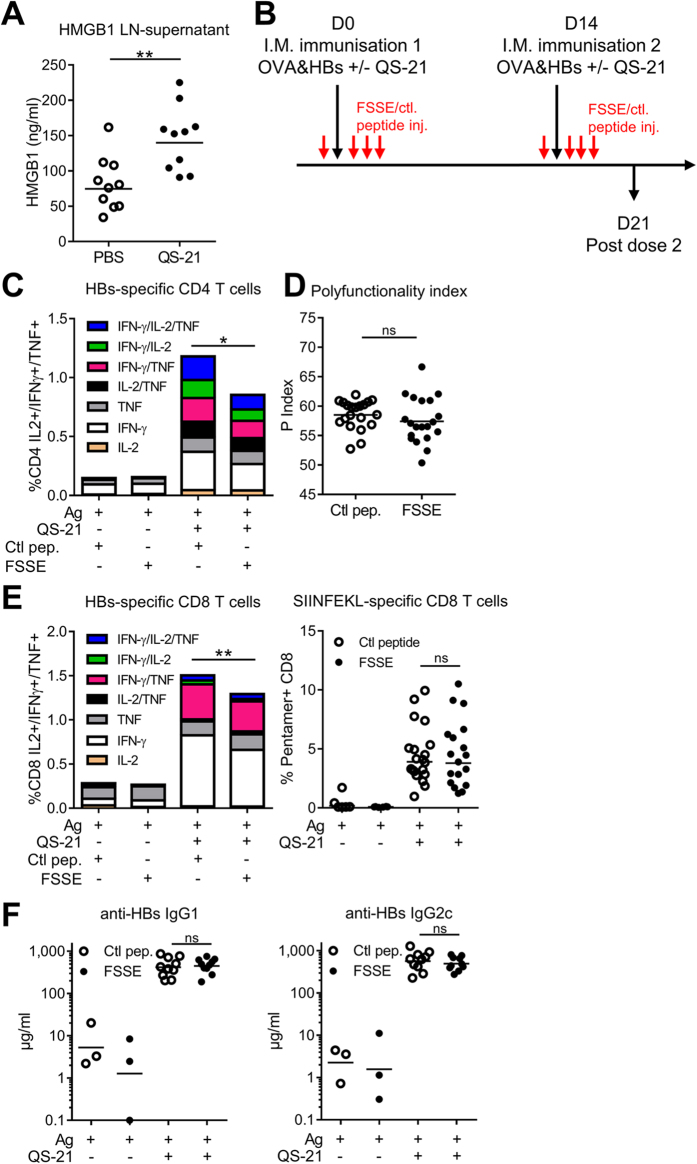
Saponins represent a promising class of vaccine adjuvant. Together with the TLR4-ligand MPL, QS-21 is part of the Adjuvant System AS01, a key component of the malaria and zoster candidate vaccines that display demonstrated clinical efficacy. However, the mechanism of action of QS-21 in this liposomal formulation is poorly understood. Upon intra-muscular immunisation, we observed that QS-21 rapidly accumulated in CD169+ resident macrophages of the draining lymph node where it elicited a local innate immune response. Depletion of these cells abrogated QS-21-mediated innate cell recruitment to the lymph node, dendritic cell (DC) phenotypic maturation as well as the adjuvant effect on T-cell and antibody responses to co-administered antigens. DCs rather than lymph node-resident macrophages were directly involved in T-cell priming by QS-21, as revealed by the decrease in antigen-specific T-cell response in Batf3-/- mice. Further analysis showed that the adjuvant effect of QS-21 depended on the integration of Caspase-1 and MyD88 pathways, at least in part through the local release of HMGB1. Taken together, this work unravels the key role of lymph node sentinel macrophage in controlling the adjuvant effect of a molecule proven to improve vaccine response in humans.
Conflict of interest statement
All authors have declared the following interests. C.C., S.W., M.C., A.E., A.M.D. are employees of the GSK group of companies. S.W., A.E. and A.M.D. own GSK. stocks. The other authors report no financial conflicts of interest.
Figures
References
-
- Reed S. G., Orr M. T. & Fox C. B. Key roles of adjuvants in modern vaccines. Nat. Med. 19, 1597–608 (2013). - PubMed
-
- Lal H. et al.. Efficacy of an adjuvanted herpes zoster subunit vaccine in older adults. N. Engl. J. Med. 372, 2087–96 (2015). - PubMed
-
- Garçon N. & Van Mechelen M. Recent clinical experience with vaccines using MPL- and QS-21-containing adjuvant systems. Expert Rev. Vaccines 10, 471–86 (2011). - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
