

Behavior-dependent activity patterns of GABAergic long-range projecting neurons in the rat hippocampus
- PMID: 27997999
- PMCID: PMC5363363
- DOI: 10.1002/hipo.22696
Behavior-dependent activity patterns of GABAergic long-range projecting neurons in the rat hippocampus
Abstract
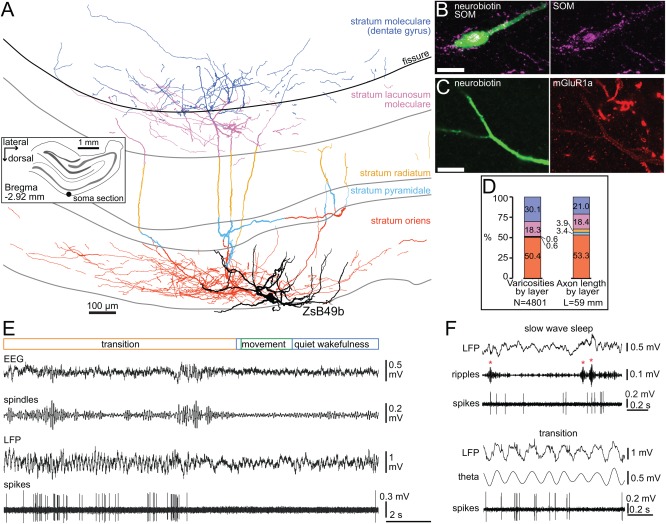
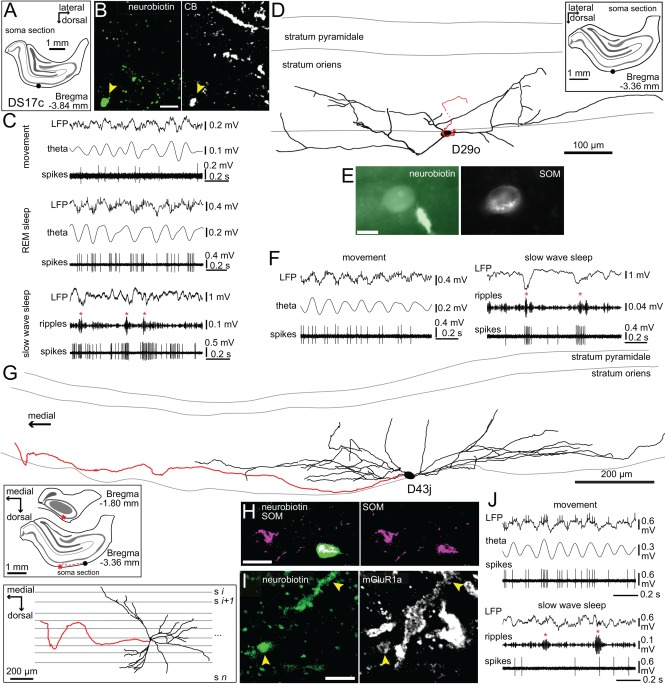
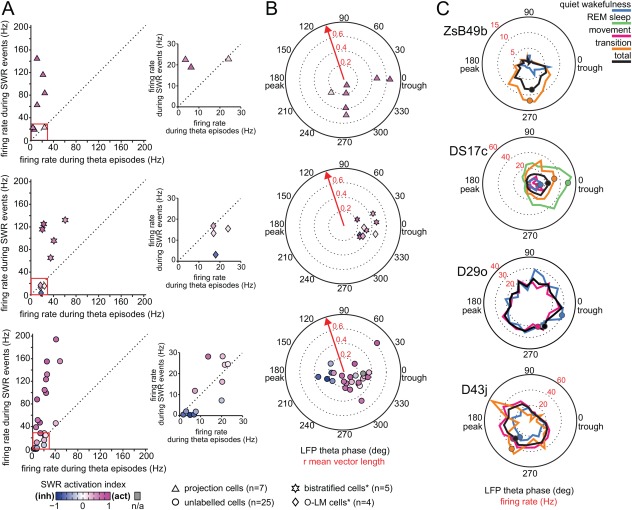
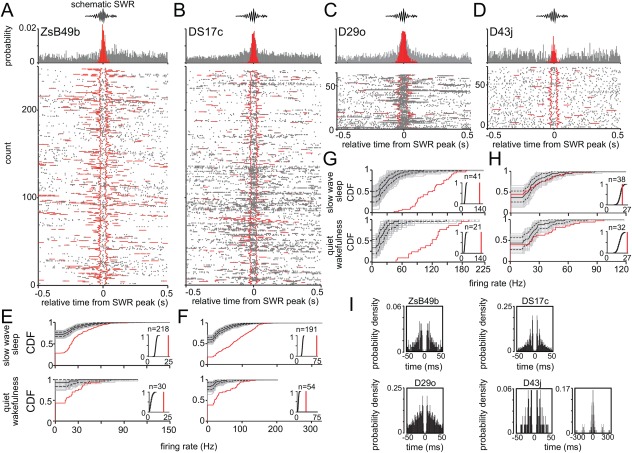
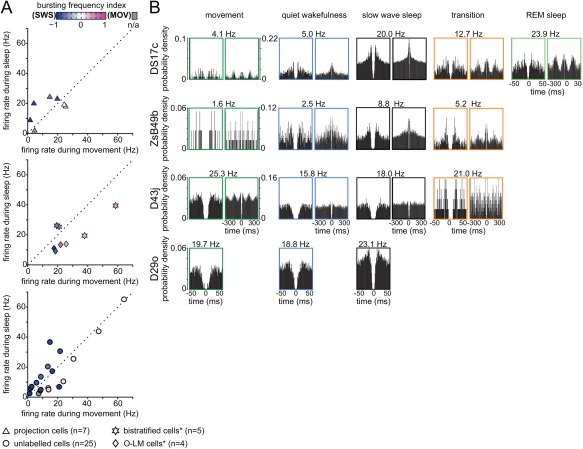
Long-range glutamatergic and GABAergic projections participate in temporal coordination of neuronal activity in distributed cortical areas. In the hippocampus, GABAergic neurons project to the medial septum and retrohippocampal areas. Many GABAergic projection cells express somatostatin (SOM+) and, together with locally terminating SOM+ bistratified and O-LM cells, contribute to dendritic inhibition of pyramidal cells. We tested the hypothesis that diversity in SOM+ cells reflects temporal specialization during behavior using extracellular single cell recording and juxtacellular neurobiotin-labeling in freely moving rats. We have demonstrated that rare GABAergic projection neurons discharge rhythmically and are remarkably diverse. During sharp wave-ripples, most projection cells, including a novel SOM+ GABAergic back-projecting cell, increased their activity similar to bistratified cells, but unlike O-LM cells. During movement, most projection cells discharged along the descending slope of theta cycles, but some fired at the trough jointly with bistratified and O-LM cells. The specialization of hippocampal SOM+ projection neurons complements the action of local interneurons in differentially phasing inputs from the CA3 area to CA1 pyramidal cell dendrites during sleep and wakefulness. Our observations suggest that GABAergic projection cells mediate the behavior- and network state-dependent binding of neuronal assemblies amongst functionally-related brain regions by transmitting local rhythmic entrainment of neurons in CA1 to neuronal populations in other areas. © 2016 The Authors Hippocampus Published by Wiley Periodicals, Inc.
Keywords: CA1; dendritic inhibition; sharp wave-ripples; somatostatin; theta oscillations.
© 2016 The Authors Hippocampus Published by Wiley Periodicals, Inc.
Figures
Similar articles
-
Spatio-temporal specialization of GABAergic septo-hippocampal neurons for rhythmic network activity.Brain Struct Funct. 2018 Jun;223(5):2409-2432. doi: 10.1007/s00429-018-1626-0. Epub 2018 Mar 3. Brain Struct Funct. 2018. PMID: 29500537 Free PMC article.
-
Neuronal diversity in GABAergic long-range projections from the hippocampus.J Neurosci. 2007 Aug 15;27(33):8790-804. doi: 10.1523/JNEUROSCI.1847-07.2007. J Neurosci. 2007. PMID: 17699661 Free PMC article.
-
Sleep and movement differentiates actions of two types of somatostatin-expressing GABAergic interneuron in rat hippocampus.Neuron. 2014 May 21;82(4):872-86. doi: 10.1016/j.neuron.2014.04.007. Epub 2014 May 1. Neuron. 2014. PMID: 24794095 Free PMC article.
-
Hippocampus as comparator: role of the two input and two output systems of the hippocampus in selection and registration of information.Hippocampus. 2001;11(5):578-98. doi: 10.1002/hipo.1073. Hippocampus. 2001. PMID: 11732710 Review.
-
Septo-hippocampal interaction.Cell Tissue Res. 2018 Sep;373(3):565-575. doi: 10.1007/s00441-017-2745-2. Epub 2017 Dec 18. Cell Tissue Res. 2018. PMID: 29250747 Free PMC article. Review.
Cited by
-
Functional specialization of hippocampal somatostatin-expressing interneurons.Proc Natl Acad Sci U S A. 2024 Apr 23;121(17):e2306382121. doi: 10.1073/pnas.2306382121. Epub 2024 Apr 19. Proc Natl Acad Sci U S A. 2024. PMID: 38640347 Free PMC article.
-
Long-Range GABAergic Projections of Cortical Origin in Brain Function.Front Syst Neurosci. 2022 Mar 22;16:841869. doi: 10.3389/fnsys.2022.841869. eCollection 2022. Front Syst Neurosci. 2022. PMID: 35392440 Free PMC article. Review.
-
Distinct Functional Classes of CA1 Hippocampal Interneurons Are Modulated by Cerebellar Stimulation in a Coordinated Manner.J Neurosci. 2024 Dec 4;44(49):e0887242024. doi: 10.1523/JNEUROSCI.0887-24.2024. J Neurosci. 2024. PMID: 39448264 Free PMC article.
-
Septo-hippocampal dynamics and the encoding of space and time.Trends Neurosci. 2023 Sep;46(9):712-725. doi: 10.1016/j.tins.2023.06.004. Epub 2023 Jul 19. Trends Neurosci. 2023. PMID: 37479632 Free PMC article. Review.
-
Behavior-Dependent Activity and Synaptic Organization of Septo-hippocampal GABAergic Neurons Selectively Targeting the Hippocampal CA3 Area.Neuron. 2017 Dec 20;96(6):1342-1357.e5. doi: 10.1016/j.neuron.2017.10.033. Epub 2017 Nov 30. Neuron. 2017. PMID: 29198757 Free PMC article.
References
-
- Acsády L, Görcs TJ, Freund TF. 1996. Different populations of vasoactive intestinal polypeptide‐immunoreactive interneurons are specialized to control pyramidal cells or interneurons in the hippocampus. Neuroscience 73:317–334. - PubMed
-
- Amaral DG, Witter MP. 1989. The three‐dimensional organization of the hippocampal formation: A review of anatomical data. Neuroscience 31:571–591. - PubMed
-
- Blasco‐Ibáñez JM, Freund TF. 1995. Synaptic input of horizontal interneurons in Stratum Oriens of the hippocampal CA1 subfield: Structural basis of feed‐back activation. Eur J Neurosci 7:2170–2180. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
