

Reprogramming of TLR7 signaling enhances antitumor NK and cytotoxic T cell responses
- PMID: 27999742
- PMCID: PMC5139637
- DOI: 10.1080/2162402X.2016.1232219
Reprogramming of TLR7 signaling enhances antitumor NK and cytotoxic T cell responses
Abstract
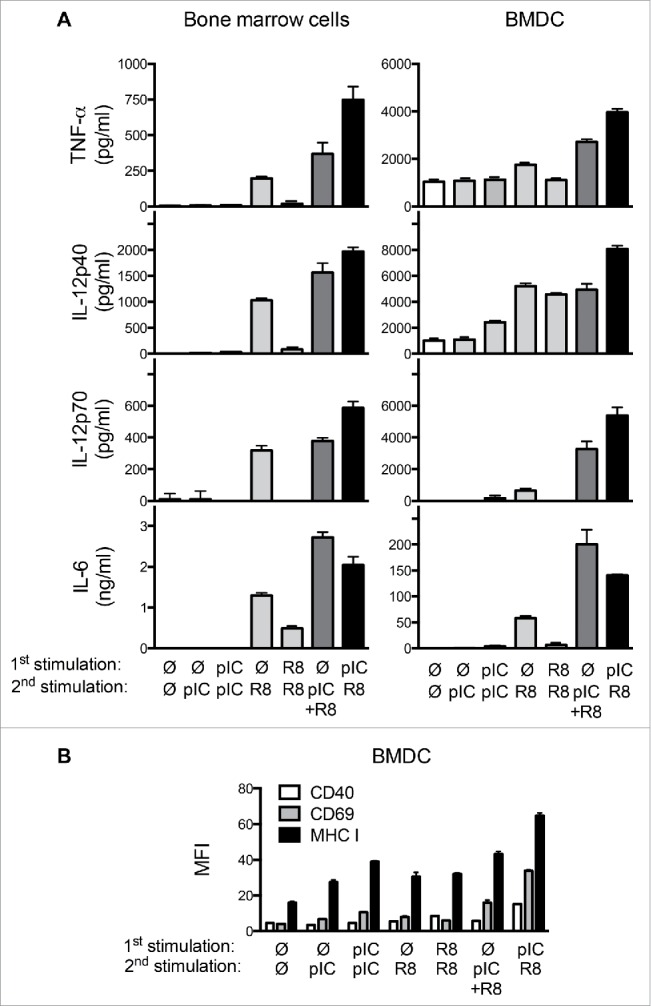
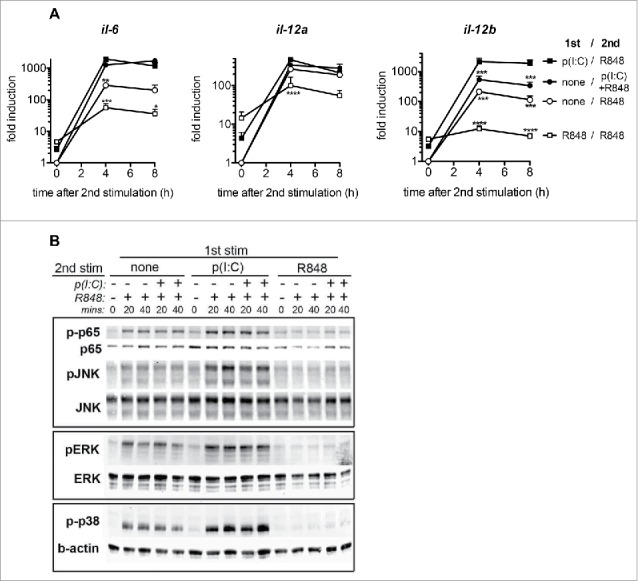
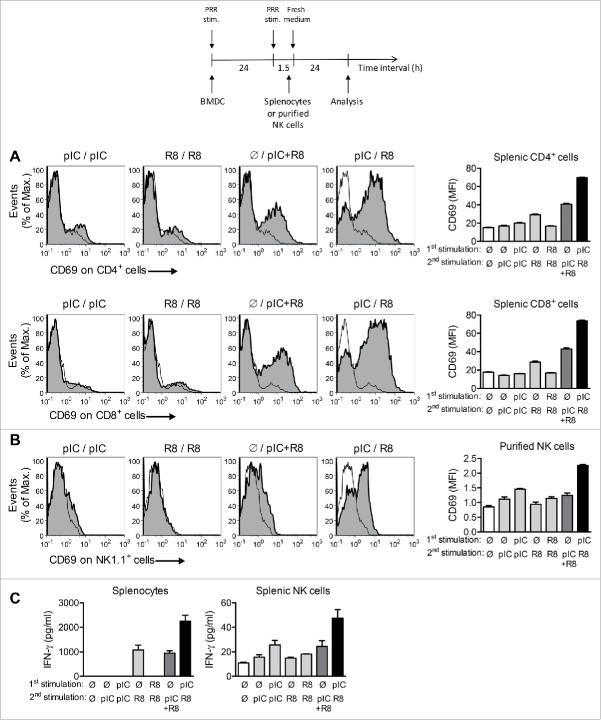
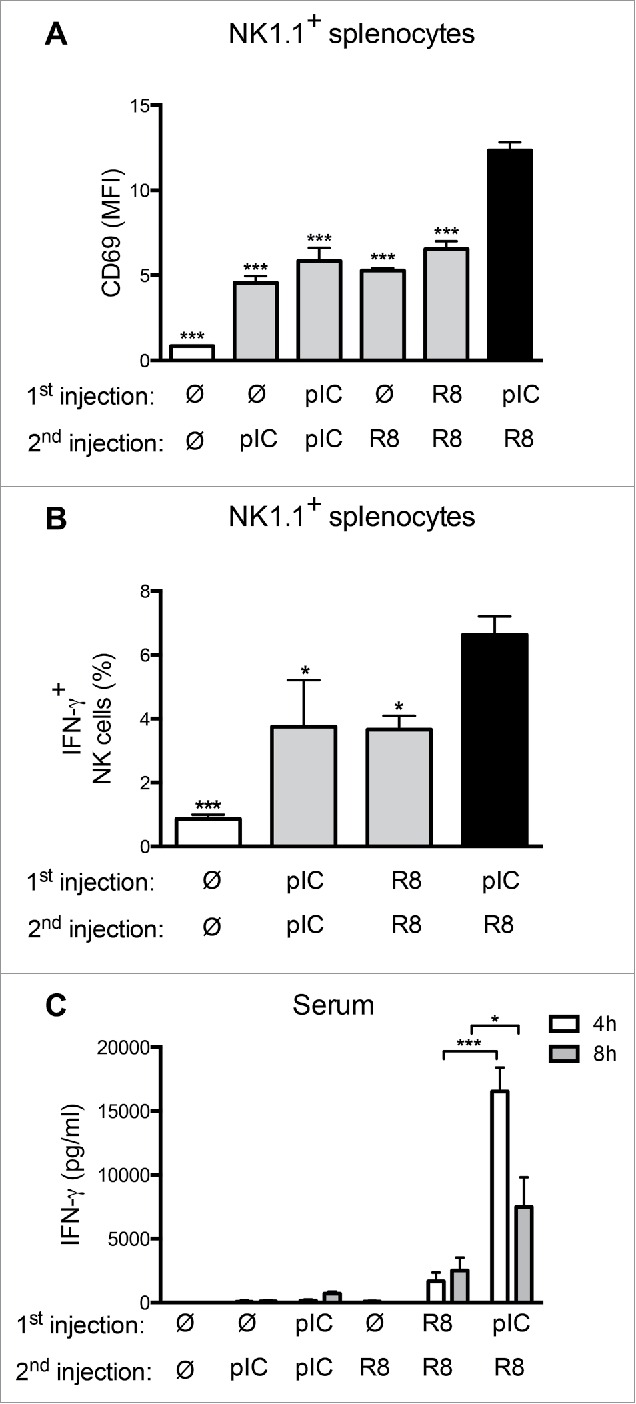
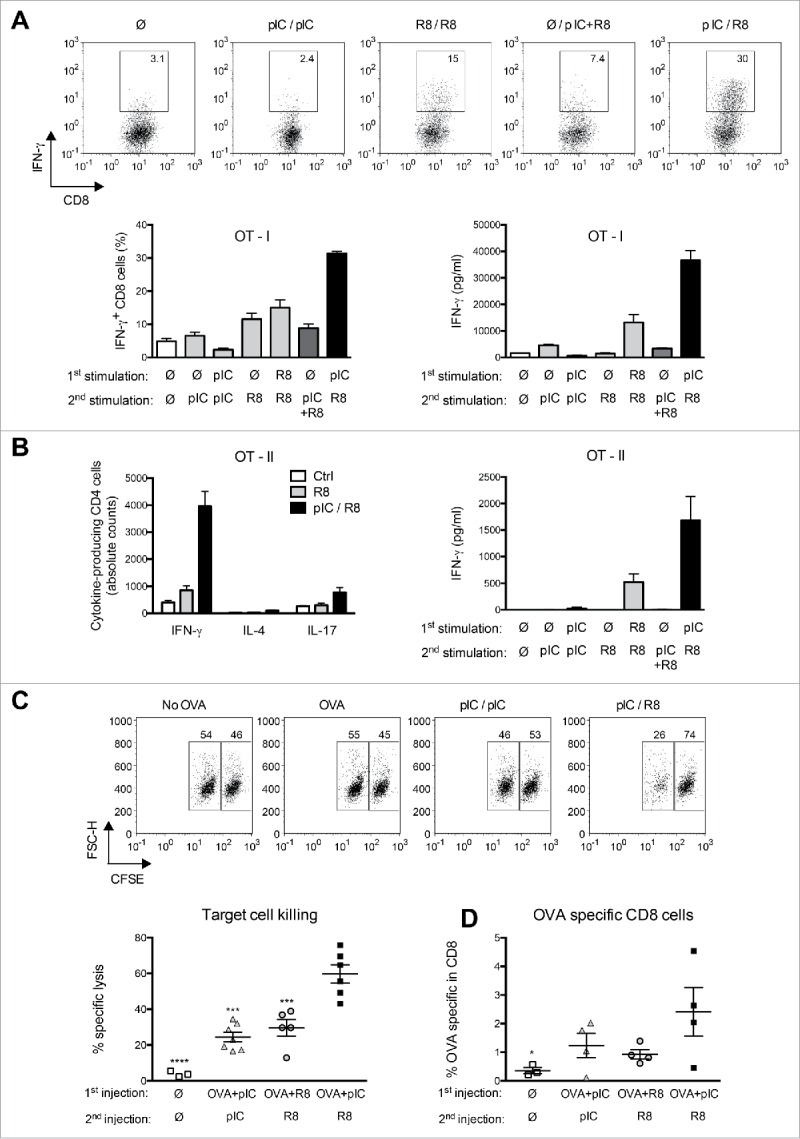
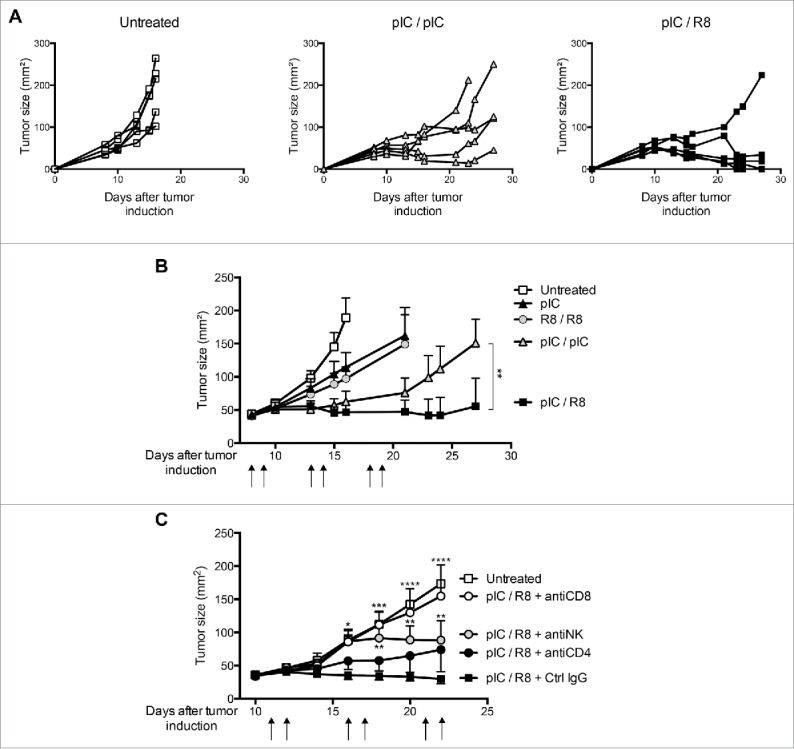
Toll-like receptor (TLR) 7 agonists are effective in topical application for the immunotherapy of skin cancers, but their performance for the systemic treatment of solid tumors is limited by the development of TLR tolerance. In this study, we describe a novel strategy to overcome TLR tolerance and enhance TLR7-dependent antitumor immune responses through reprogramming of TLR signaling pathways. The sensitivity of TLR7 signaling in dendritic cells (DC) was increased by prior stimulation with the dsRNA poly(I:C) that mimics virally induced immune activation. Timing of the stimulations was important, as sequential stimulation with poly(I:C) and the TLR7 agonist R848 interspaced by 24 h induced higher MAPK and NFkB signaling in DC than the simultaneous application of the same ligands. DC activated by sequential poly(I:C)/R848 stimulation efficiently induced Th1 differentiation and primed NK-cell and cytotoxic T-cell responses. We have developed a treatment regimen taking advantage of TLR7 reprogram-ming that cured over 80% of large immunogenic tumors in mice by the action of NK cells and cytotoxic T cells. These results have direct implications for the use of these clinically established ligands in the immunotherapy of cancer.
Keywords: Cancer immunotherapy; RIG-I-like receptors agonists; Toll-like receptors; dendritic cells; poly(I:C).
Figures
References
-
- Makkouk A, Weiner GJ. Cancer immunotherapy and breaking immune tolerance: new approaches to an old challenge. Cancer Res 2015; 75:5-10; PMID:25524899; http://dx.doi.org/ 10.1158/0008-5472.CAN-14-2538 - DOI - PMC - PubMed
-
- Steinman RM, Hemmi H. Dendritic cells: translating innate to adaptive immunity. Curr Top Microbiol Immunol 2006; 311:17-58; PMID:17048704; http://dx.doi.org/ 10.1007/3-540-32636-7_2 - DOI - PubMed
-
- Lucas M, Schachterle W, Oberle K, Aichele P, Diefenbach A. Dendritic cells prime natural killer cells by trans-presenting interleukin 15. Immunity 2007; 26:503-17; PMID:17398124; http://dx.doi.org/ 10.1016/j.immuni.2007.03.006 - DOI - PMC - PubMed
-
- Junt T, Barchet W. Translating nucleic acid-sensing pathways into therapies. Nat Rev Immunol 2015; 15:529-44; PMID:26292638; http://dx.doi.org/ 10.1038/nri3875 - DOI - PubMed
-
- Peris K, Campione E, Micantonio T, Marulli GC, Fargnoli MC, Chimenti S. Imiquimod treatment of superficial and nodular basal cell carcinoma: 12-week open-label trial. Dermatol Surg Off Publ Am Soc Dermatol Surg Al 2005; 31:318-23; PMID:15841634 - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources