

Hematopoietic Stem Cells in Neural-crest Derived Bone Marrow
- PMID: 28000662
- PMCID: PMC5175267
- DOI: 10.1038/srep36411
Hematopoietic Stem Cells in Neural-crest Derived Bone Marrow
Abstract
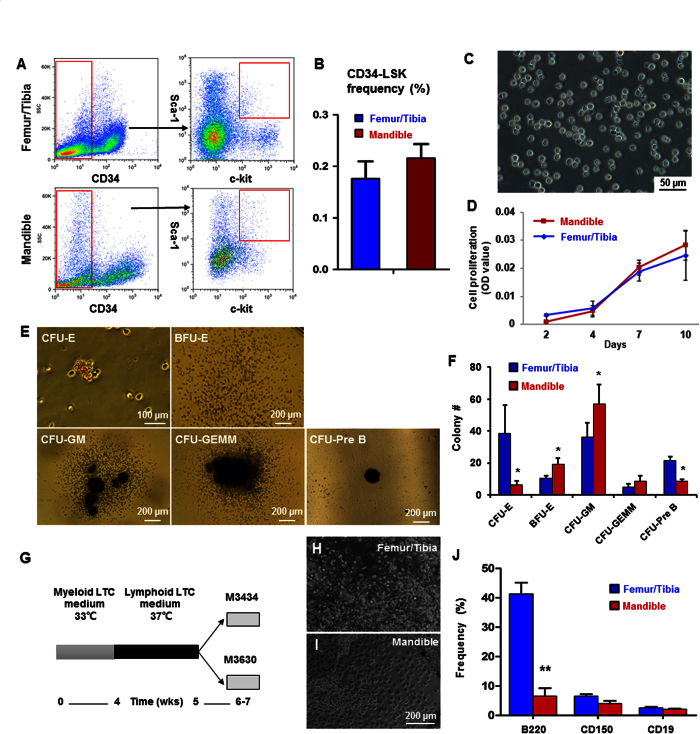
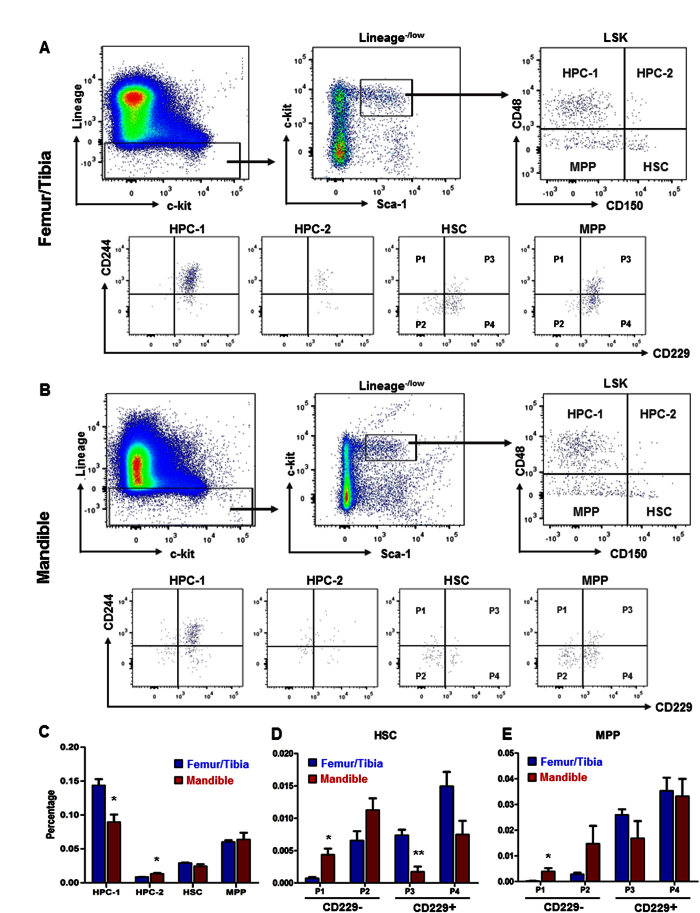
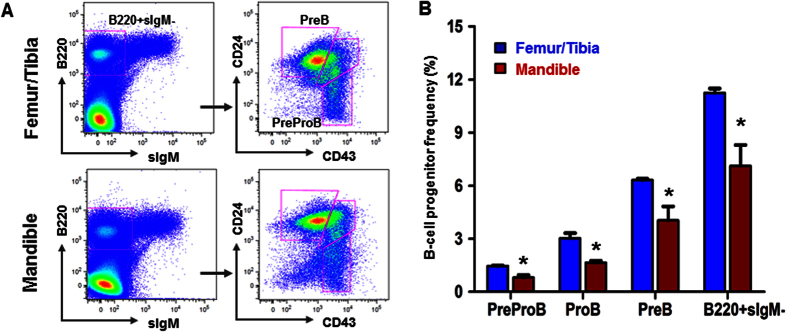
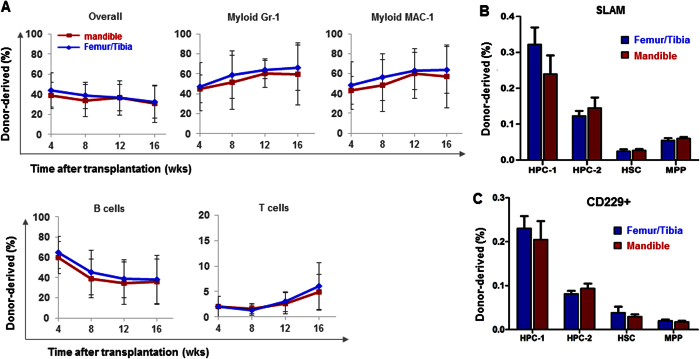
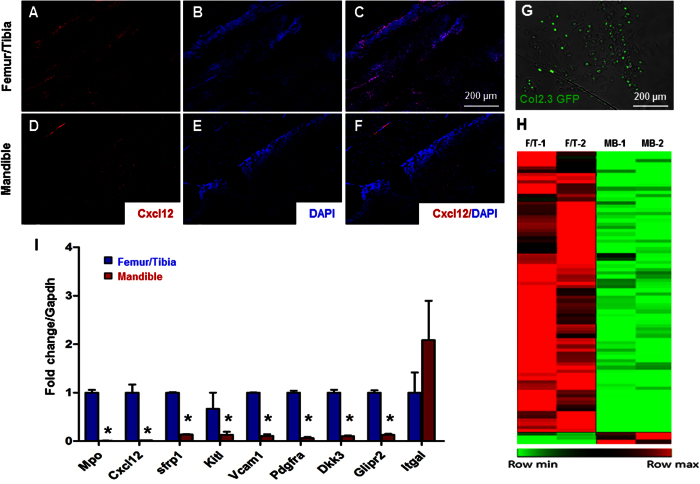
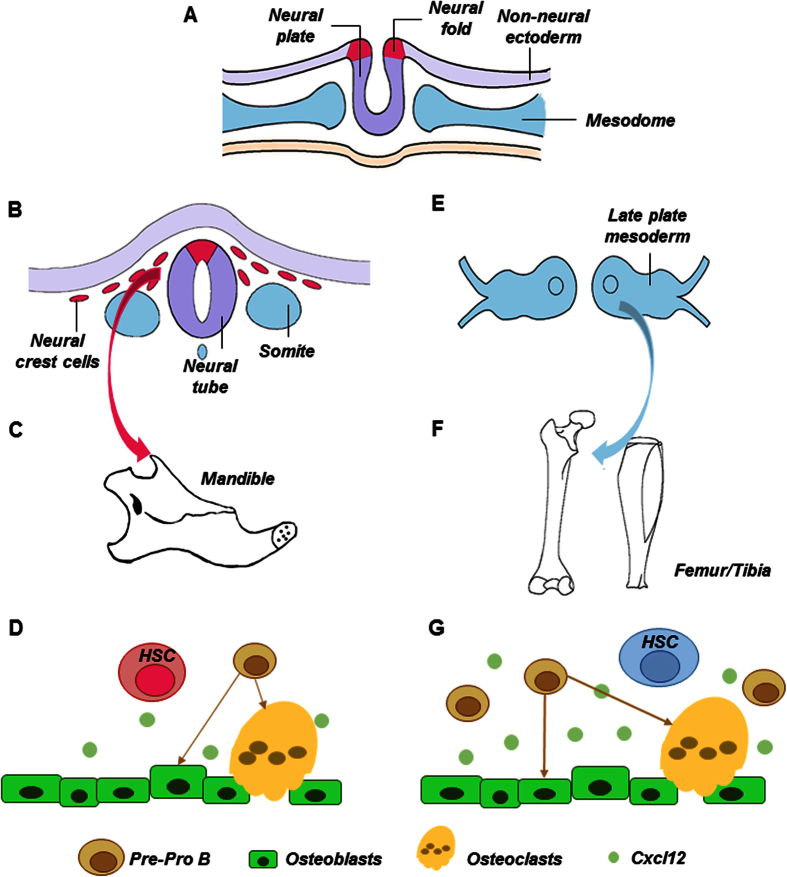
Hematopoietic stem cells (HSCs) in the endosteum of mesoderm-derived appendicular bones have been extensively studied. Neural crest-derived bones differ from appendicular bones in developmental origin, mode of bone formation and pathological bone resorption. Whether neural crest-derived bones harbor HSCs is elusive. Here, we discovered HSC-like cells in postnatal murine mandible, and benchmarked them with donor-matched, mesoderm-derived femur/tibia HSCs, including clonogenic assay and long-term culture. Mandibular CD34 negative, LSK cells proliferated similarly to appendicular HSCs, and differentiated into all hematopoietic lineages. Mandibular HSCs showed a consistent deficiency in lymphoid differentiation, including significantly fewer CD229 + fractions, PreProB, ProB, PreB and B220 + slgM cells. Remarkably, mandibular HSCs reconstituted irradiated hematopoietic bone marrow in vivo, just as appendicular HSCs. Genomic profiling of osteoblasts from mandibular and femur/tibia bone marrow revealed deficiencies in several HSC niche regulators among mandibular osteoblasts including Cxcl12. Neural crest derived bone harbors HSCs that function similarly to appendicular HSCs but are deficient in the lymphoid lineage. Thus, lymphoid deficiency of mandibular HSCs may be accounted by putative niche regulating genes. HSCs in craniofacial bones have functional implications in homeostasis, osteoclastogenesis, immune functions, tumor metastasis and infections such as osteonecrosis of the jaw.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Spangrude G. J., Heimfeld S. & Weissman I. L. Purification and characterization of mouse hematopoietic stem cells. Science 241, 58–62 (1988). - PubMed
-
- Warr M. R., Pietras E. M. & Passegue E. Mechanisms controlling hematopoietic stem cell functions during normal hematopoiesis and hematological malignancies. Wiley interdisciplinary reviews. Systems biology and medicine 3, 681–701 (2011). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
