

Piezo2 senses airway stretch and mediates lung inflation-induced apnoea
- PMID: 28002412
- PMCID: PMC5267560
- DOI: 10.1038/nature20793
Piezo2 senses airway stretch and mediates lung inflation-induced apnoea
Abstract
Respiratory dysfunction is a notorious cause of perinatal mortality in infants and sleep apnoea in adults, but the mechanisms of respiratory control are not clearly understood. Mechanical signals transduced by airway-innervating sensory neurons control respiration; however, the physiological significance and molecular mechanisms of these signals remain obscured. Here we show that global and sensory neuron-specific ablation of the mechanically activated ion channel Piezo2 causes respiratory distress and death in newborn mice. Optogenetic activation of Piezo2+ vagal sensory neurons causes apnoea in adult mice. Moreover, induced ablation of Piezo2 in sensory neurons of adult mice causes decreased neuronal responses to lung inflation, an impaired Hering-Breuer mechanoreflex, and increased tidal volume under normal conditions. These phenotypes are reproduced in mice lacking Piezo2 in the nodose ganglion. Our data suggest that Piezo2 is an airway stretch sensor and that Piezo2-mediated mechanotransduction within various airway-innervating sensory neurons is critical for establishing efficient respiration at birth and maintaining normal breathing in adults.
Figures

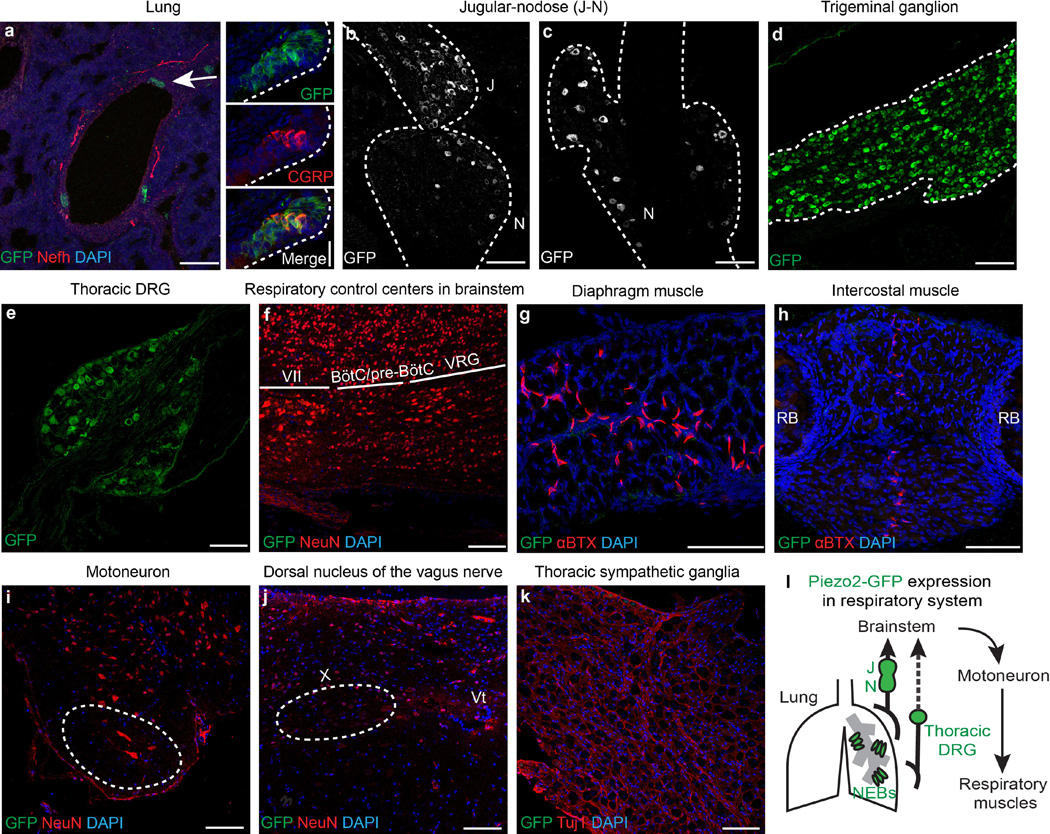
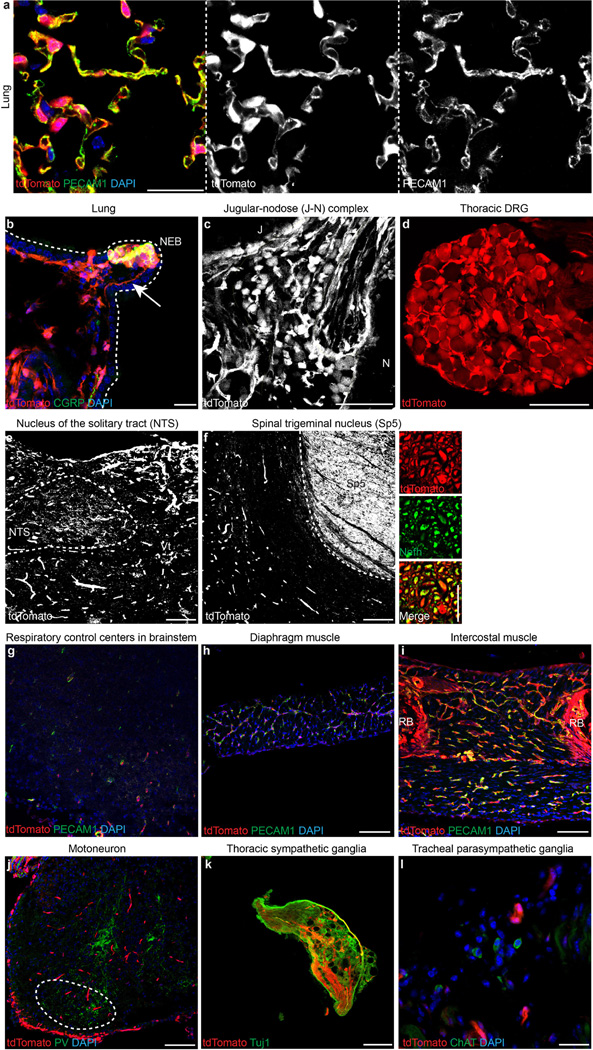


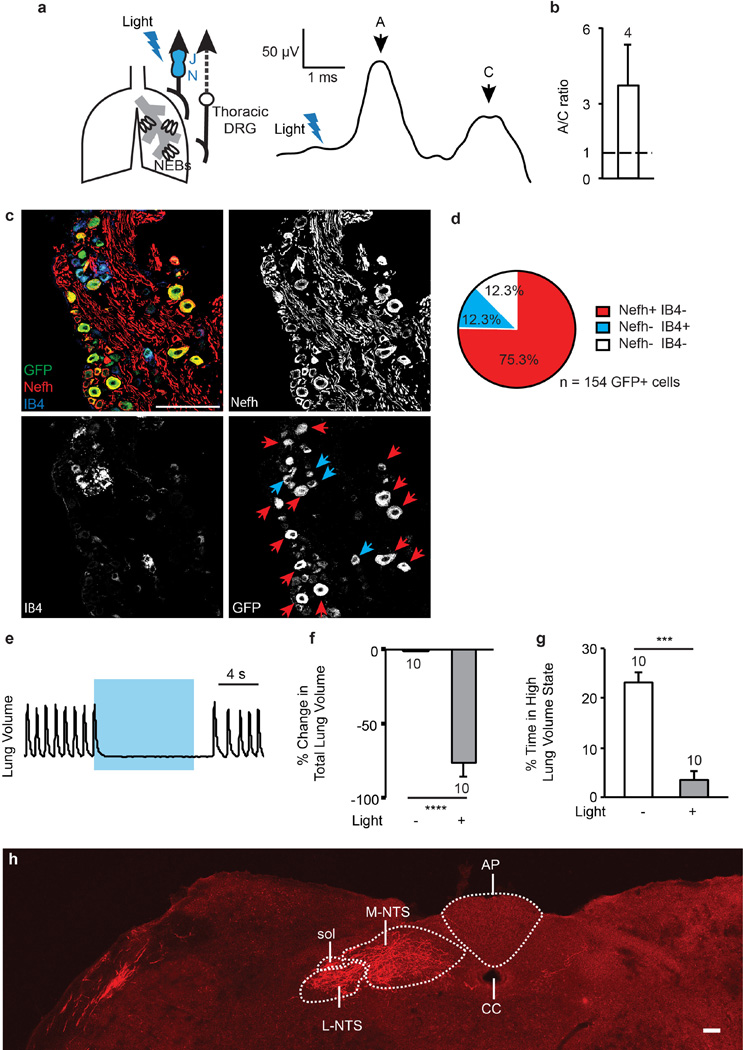








Comment in
-
Physiology: Mechanosensor of lung inflation identified.Nature. 2017 Jan 12;541(7636):165-166. doi: 10.1038/nature21111. Epub 2016 Dec 21. Nature. 2017. PMID: 28002406 No abstract available.
References
-
- Rhoades RA, Bell DR. Medical Physiology: Principles for Clinical Medicine. 3rd. Vol. 328. Lippincott Williams & Wilkins; 2009.
-
- Schelegle ES, Green JF. An overview of the anatomy and physiology of slowly adapting pulmonary stretch receptors. Respir. Physiol. 2001;125:17–31. - PubMed
-
- Lee LY, Yu J. Sensory nerves in lung and airways. Compr. Physiol. 2014;4:287–324. - PubMed
-
- Zhang JW, Walker JF, Guardiola J, Yu J. Pulmonary sensory and reflex responses in the mouse. J. Appl. Physiol. 2006;101:986–992. - PubMed
-
- Carr MJ, Undem BJ. Bronchopulmonary afferent nerves. Respirology. 2003;8:291–301. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
