

Shifting Clade Distribution, Reassortment, and Emergence of New Subtypes of Highly Pathogenic Avian Influenza A(H5) Viruses Collected from Vietnamese Poultry from 2012 to 2015
- PMID: 28003481
- PMCID: PMC5309939
- DOI: 10.1128/JVI.01708-16
Shifting Clade Distribution, Reassortment, and Emergence of New Subtypes of Highly Pathogenic Avian Influenza A(H5) Viruses Collected from Vietnamese Poultry from 2012 to 2015
Abstract
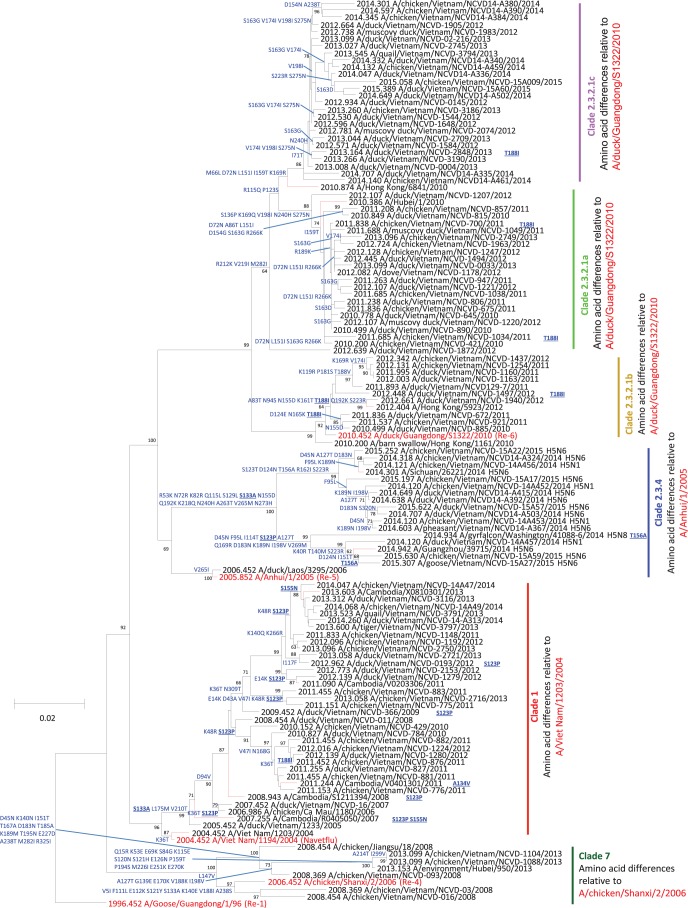
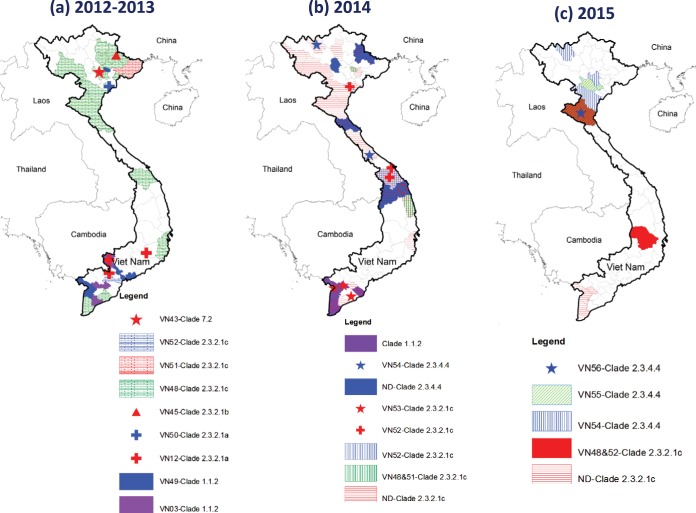
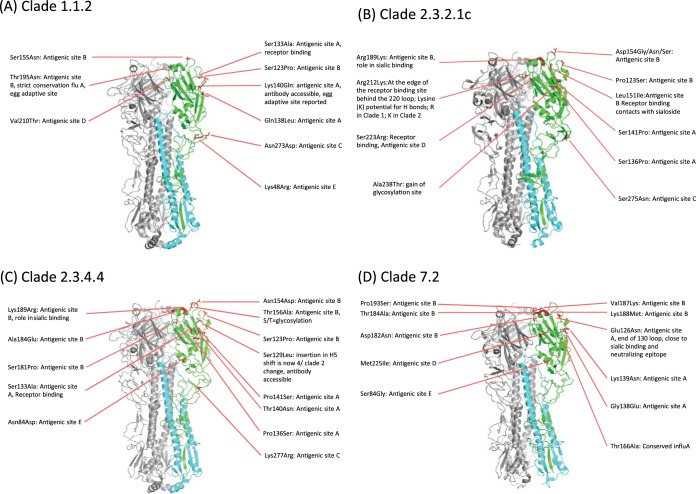
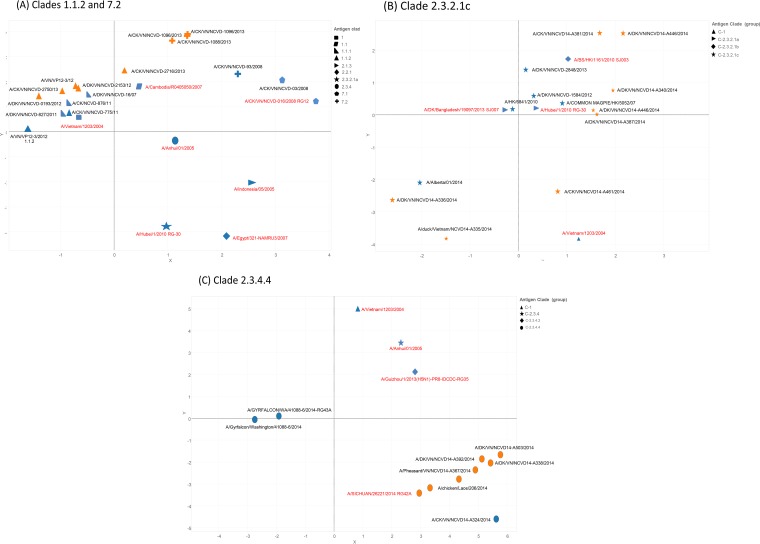
Whole-genome sequences of representative highly pathogenic avian influenza A(H5) viruses from Vietnam were generated, comprising samples from poultry outbreaks and active market surveillance collected from January 2012 to August 2015. Six hemagglutinin gene clades were characterized. Clade 1.1.2 was predominant in southern Mekong provinces throughout 2012 and 2013 but gradually disappeared and was not detected after April 2014. Clade 2.3.2.1c viruses spread rapidly during 2012 and were detected in the south and center of the country. A number of clade 1.1.2 and 2.3.2.1c interclade reassortant viruses were detected with different combinations of internal genes derived from 2.3.2.1a and 2.3.2.1b viruses, indicating extensive cocirculation. Although reassortment generated genetic diversity at the genotype level, there was relatively little genetic drift within the individual gene segments, suggesting genetic stasis over recent years. Antigenically, clade 1.1.2, 2.3.2.1a, 2.3.2.1b, and 2.3.2.1c viruses remained related to earlier viruses and WHO-recommended prepandemic vaccine strains representing these clades. Clade 7.2 viruses, although detected in only low numbers, were the exception, as indicated by introduction of a genetically and antigenically diverse strain in 2013. Clade 2.3.4.4 viruses (H5N1 and H5N6) were likely introduced in April 2014 and appeared to gain dominance across northern and central regions. Antigenic analyses of clade 2.3.4.4 viruses compared to existing clade 2.3.4 candidate vaccine viruses (CVV) indicated the need for an updated vaccine virus. A/Sichuan/26221/2014 (H5N6) virus was developed, and ferret antisera generated against this virus were demonstrated to inhibit some but not all clade 2.3.4.4 viruses, suggesting consideration of alternative clade 2.3.4.4 CVVs.IMPORTANCE Highly pathogenic avian influenza (HPAI) A(H5) viruses have circulated continuously in Vietnam since 2003, resulting in hundreds of poultry outbreaks and sporadic human infections. Despite a significant reduction in the number of human infections in recent years, poultry outbreaks continue to occur and the virus continues to diversify. Vaccination of poultry has been used as a means to control the spread and impact of the virus, but due to the diversity and changing distribution of antigenically distinct viruses, the utility of vaccines in the face of mismatched circulating strains remains questionable. This study assessed the putative amino acid changes in viruses leading to antigenic variability, underscoring the complexity of vaccine selection for both veterinary and public health purposes. Given the overlapping geographic distributions of multiple, antigenically distinct clades of HPAI A(H5) viruses in Vietnam, the vaccine efficacy of bivalent poultry vaccine formulations should be tested in the future.
Keywords: Vietnam; antigenicity; evolution; highly pathogenic avian influenza; reassortment; vaccine selection.
Copyright © 2017 American Society for Microbiology.
Figures

References
-
- Creanga A, Diep Thi N, Gerloff N, Hoa Thi D, Balish A, Hoang Dang N, Jang Y, Vui Thi D, Thor S, Jones J, Simpson N, Shu B, Emery S, Berman L, Nguyen HT, Bryant JE, Lindstrom S, Klimov A, Donis RO, Davis CT, Tung N. 2013. Emergence of multiple clade 2.3.2.1 influenza A (H5N1) virus subgroups in Vietnam and detection of novel reassortants. Virology 444:12–20. doi: 10.1016/j.virol.2013.06.005. - DOI - PubMed
-
- Nguyen T, Rivailler P, Davis CT, Hoa do T, Balish A, Dang NH, Jones J, Vui DT, Simpson N, Huong NT, Shu B, Loughlin R, Ferdinand K, Lindstrom SE, York IA, Klimov A, Donis RO. 2012. Evolution of highly pathogenic avian influenza (H5N1) virus populations in Vietnam between 2007 and 2010. Virology 432:405–416. doi: 10.1016/j.virol.2012.06.021. - DOI - PubMed
-
- Wan XF, Nguyen T, Davis CT, Smith CB, Zhao ZM, Carrel M, Inui K, Do HT, Mai DT, Jadhao S, Balish A, Shu B, Luo F, Emch M, Matsuoka Y, Lindstrom SE, Cox NJ, Nguyen CV, Klimov A, Donis RO. 21 October 2008. Evolution of highly pathogenic H5N1 avian influenza viruses in Viet Nam between 2001 and 2007. PLoS One doi: 10.1371/journal.pone.0003462. - DOI - PMC - PubMed
-
- Rith S, Davis CT, Duong V, Sar B, Horm SV, Chin S, Ly S, Laurent D, Richner B, Oboho I, Jang Y, Davis W, Thor S, Balish A, Iuliano AD, Sorn S, Holl D, Sok T, Seng H, Tarantola A, Tsuyuoka R, Parry A, Chea N, Allal L, Kitsutani P, Warren D, Prouty M, Horwood P, Widdowson MA, Lindstrom S, Villanueva J, Donis R, Cox N, Buchy P. 10 September 2014. Identification of molecular markers associated with alteration of receptor-binding specificity in a novel genotype of highly pathogenic avian influenza A (H5N1) viruses detected in Cambodia in 2013. J Virol doi: 10.1128/JVI.01887-14. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
