

doi: 10.1038/ncomms13721.
Top-down modulation of olfactory-guided behaviours by the anterior olfactory nucleus pars medialis and ventral hippocampus
Affiliations
- PMID: 28004701
- PMCID: PMC5192165
- DOI: 10.1038/ncomms13721
Item in Clipboard
Top-down modulation of olfactory-guided behaviours by the anterior olfactory nucleus pars medialis and ventral hippocampus
Nat Commun.
.
Abstract
Olfactory processing is thought to be actively modulated by the top-down input from cortical regions, but the behavioural function of these signals remains unclear. Here we find that cortical feedback from the anterior olfactory nucleus pars medialis (mAON) bidirectionally modulates olfactory sensitivity and olfaction-dependent behaviours. To identify a limbic input that tunes this mAON switch, we further demonstrate that optogenetic stimulation of ventral hippocampal inputs to the mAON is sufficient to alter olfaction-dependent behaviours.
Figures
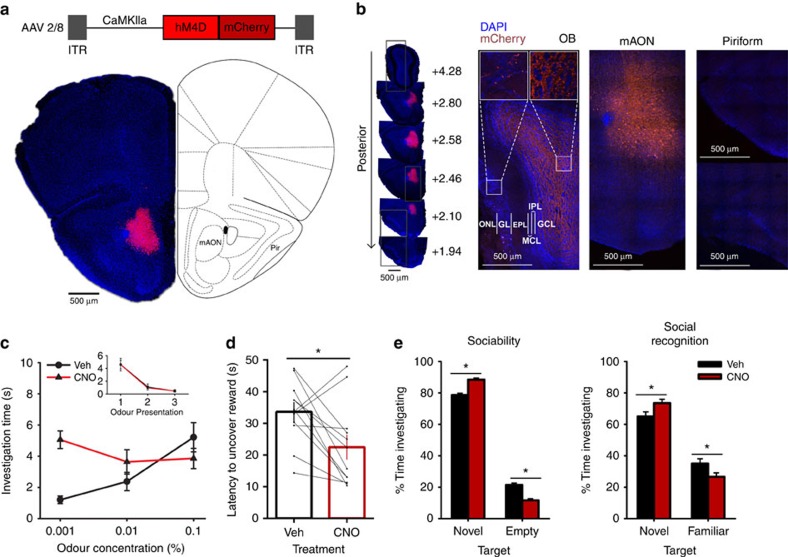
(a) AAV-mediated expression of hM4D-mCherry in CaMKIIa-positive neurons was restricted to the mAON. ITR, inverted terminal repeats. (b, left) Serial sections depicting the extent of hM4D-mCherry expression at viral infusion site with AP axis coordinates from bregma for reference. (right) Confocal images of OB coronal sections depicting DAPI-stained nuclei and mCherry-positive axon fibres arriving from mAON. Right panels correspond to the boxed regions in the left panel. mCherry-positive axon terminals of mAON CaMKIIa-positive neurons were found to innervate primarily the deep granule cell layer with a few scattered fibres present in the glomerular layer of the OB (EPL, external plexiform layer; GCL, granule cell layer; GL, glomerular layer; IPL, internal plexiform layer; MCL, mitral cell layer; ONL, olfactory nerve layer), but virtually none are present in the anterior (above) or posterior piriform cortex (below). (c) CNO treatment did not alter investigation of mineral oil (0% odour concentration) across habituation trials (inset; data obtained from olfactory habituation/dishabituation test, Supplementary Fig. 2a), but increased investigation time of an odour at a low concentration compared with vehicle treatment (n=11, two-way ANOVA interaction F(2,20)=10.94, *P<0.001). (d) CNO treatment decreased the latency to locate a buried food reward (n=12, paired-samples t-test, t(11)=2.85, *P<0.05). (e) Compared with vehicle-treated mice, CNO-treated mice spent proportionally more time investigating a conspecific versus an empty cage (n=11 per group, independent-samples t-test, t(20)=6.32, *P<0.001; absolute investigation time for novel conspecific between groups, t(20)=2.15, *P<0.05), and showed a higher preference for investigating a novel versus familiar conspecific (t(20)=2.18, *P<0.05; absolute investigation time for novel conspecific between groups, t(20)=6.27, *P<0.05). ANOVA, analysis of variance.
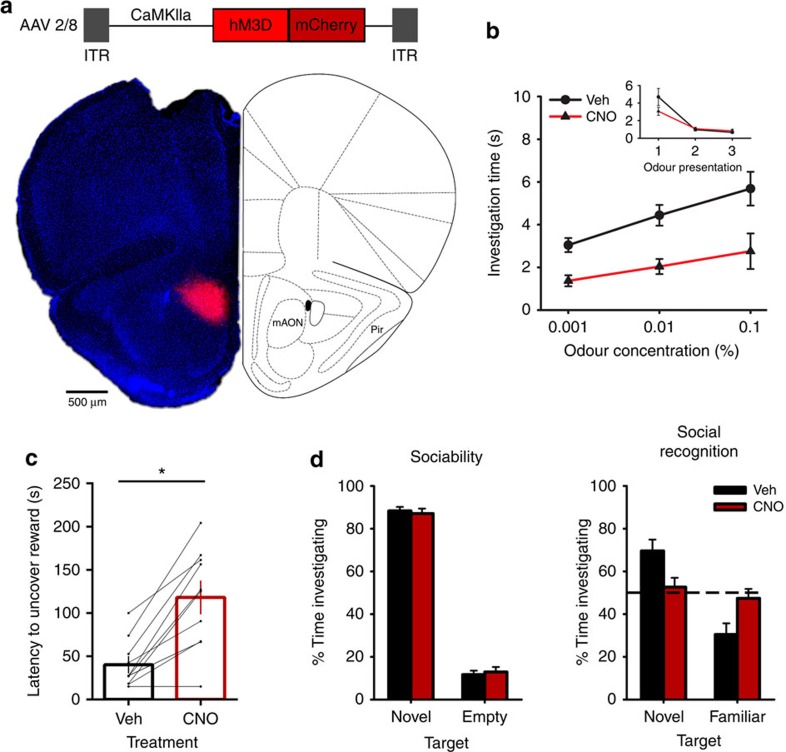
(a) AAV-mediated expression of hM3D-mCherry in CaMKIIa-positive neurons was restricted to the mAON. (b) CNO treatment did not alter investigation of mineral oil (0% odour concentration) across habituation trials (inset; data obtained from olfactory habituation/dishabituation test, Supplementary Fig. 4a), but impaired detection of an odour at a low concentration compared with vehicle treatment (n=12, two-way ANOVA both main effects F(2,22)>10.50, P<0.001). (c) CNO treatment increased the latency to locate a buried food reward (n=10, paired-samples t-test, t(9)=5.57 *P<0.001). (d) CNO-treated mice exhibited no change in sociability (vehicle group n=10, CNO group n=11, independent-samples t-test, t(19)=0.41, ns, P=0.68), but failed to distinguish between a novel and familiar conspecific (independent-samples t(19)=2.50, P<0.05; paired-samples t-tests, vehicle group t(9)=3.71, P<0.01, CNO group t(10)=0.60, ns, P=0.56; absolute investigation time for novel conspecific between groups, t(19)=1.13, ns, P=0.25). ANOVA, analysis of variance.
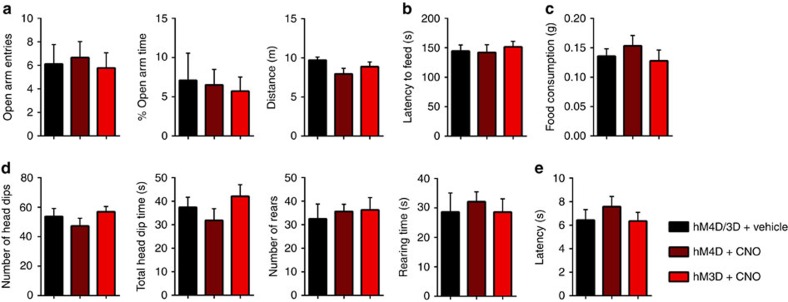
hM3D-expressing mice and hM4D-expressing mice were each divided into two groups and treated with either vehicle or CNO before the behavioural tests. Vehicle-treated hM3D mice and hM4D mice showed no difference and, therefore, were combined. (a) In an elevated-plus maze, CNO-treatment in hM4D or hM3D-expressing animals did not influence entries into the open arms (left: independent-samples t-test; vehicle (n=9) versus hM4D (n=9), t(16)=0.26, ns, P=0.80; vehicle versus hM3D (n=9), t(16)=0.16, ns, P=0.88), percentage of time spent in the open arms (middle: vehicle versus hM4D, t(16)=0.39, ns, P=0.70; vehicle versus hM3D, t(16)=0.62, ns, P=0.54), or total distance travelled (right: vehicle versus hM4D t(16)=1.85, ns, P=0.084; vehicle versus hM3D, t(16)=1.19, ns, P=0.25). (b) CNO-treated groups displayed similar latency to retrieve and consume food placed in the centre of a brightly lit open field compared with vehicle-treated groups (vehicle (n=9) versus hM4D (n=9), t(16)=0.14, ns, P=0.89; vehicle versus hM3D (n=9), t(16)=0.52, ns, P=0.61). (c) CNO-treatment did not change the amount of food consumed in the home cage (vehicle (n=9) versus hM4D (n=9), t(16)=0.82, ns, P=0.43; vehicle versus hM3D (n=9), t(16)=0.35, ns, P=0.73). (d) Exploratory behaviour in the hole-board test was not affected by inhibition or activation of the mAON as determined by measuring number of head dips (left: vehicle (n=8) versus hM4D (n=8), t(14)=0.87, ns, P=0.40; vehicle versus hM3D (n=8), t(14)=0.48, ns, P=0.64), total head dip time (middle-left: vehicle versus hM4D, t(14)=0.85, ns, P=0.41; vehicle versus hM3D, t(14)=0.71, ns, P=0.49), number of rears (middle-right: vehicle versus hM4D, t(14)=0.45, ns, P=0.66; vehicle versus hM3D, t(14)=0.46, ns, P=0.65), and right: total rearing time (vehicle versus hM4D, t(14)=0.48, ns, P=0.64; vehicle versus hM3D, t(14)=0.0056, ns, P=0.996). (e) CNO-treated hM4D and hM3D groups exhibited no difference in latency to obtain food on a runway compared with vehicle-treated group (vehicle (n=8) versus hM4D (n=8), t(14)=0.92, ns, P=0.37; vehicle versus hM3D (n=8), t(14)=0.064, ns, P=0.95).
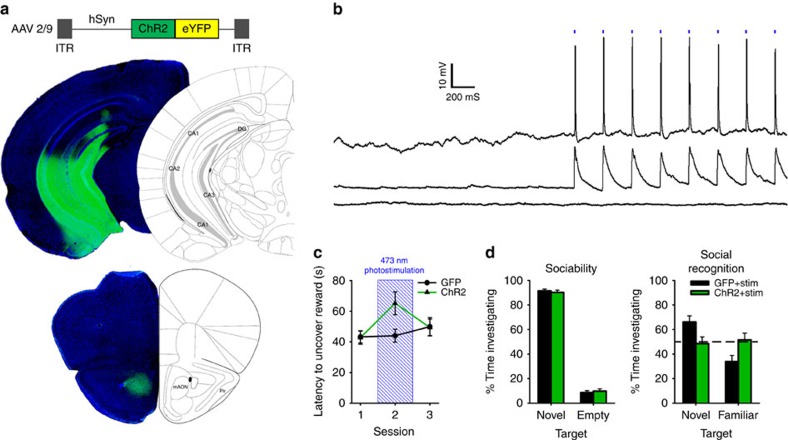
(a) AAV-mediated expression of ChR2-YFP in the vHPC driven by the human synapsin promoter (hSyn) produced dense terminal fields localized to the mAON. (b) Example traces from in vitro current-clamped mAON neurons in response to stimulation of ChR2-containing vHPC terminals (5 ms pulses, 4 Hz, 5 mW). Cells showed reliable spiking activity (n=1) or EPSPs (n=6) following light pulses, while some exhibited no response (n=2). (c) In vivo stimulation of the vHPC-mAON pathway (5 ms pulses, 4 Hz, 1 mW) significantly increased the latency to locate a buried food reward compared with control mice (n=8 per group, two-way ANOVA interaction F(2,28)=3.60, P<0.05; session 2, t(14)=2.47, P<0.05). (d) In the absence of photostimulation, ChR2 and GFP mice showed no difference in the proportion of time spent investigating a novel conspecific versus an empty cage (left: ChR2 n=7, GFP n=8; independent-samples t-test, t(13)=0.48, P=0.64). However, photostimulation impaired the ability of ChR2 mice to distinguish between a novel and familiar conspecific (right: independent-samples t-test, t(13)=2.38, P<0.05; paired-samples t-tests, GFP group t(7)=3.25, P<0.01, ChR2 group t(6)=0.60, ns; absolute investigation time for novel conspecific between groups, t(13)=0.28, ns). ANOVA, analysis of variance.
References
-
- Shepherd G. M. Synaptic organization of the mammalian olfactory bulb. Physiol. Rev. 52, 864–917 (1972). - PubMed
-
- Shipley M. T. & Adamek G. D. The connections of the mouse olfactory bulb: a study using orthograde and retrograde transport of wheat germ agglutinin conjugated to horseradish peroxidase. Brain Res. Bull 12, 669–688 (1984). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
