

Angiopoietin receptor Tie2 is required for vein specification and maintenance via regulating COUP-TFII
- PMID: 28005008
- PMCID: PMC5218530
- DOI: 10.7554/eLife.21032
Angiopoietin receptor Tie2 is required for vein specification and maintenance via regulating COUP-TFII
Abstract
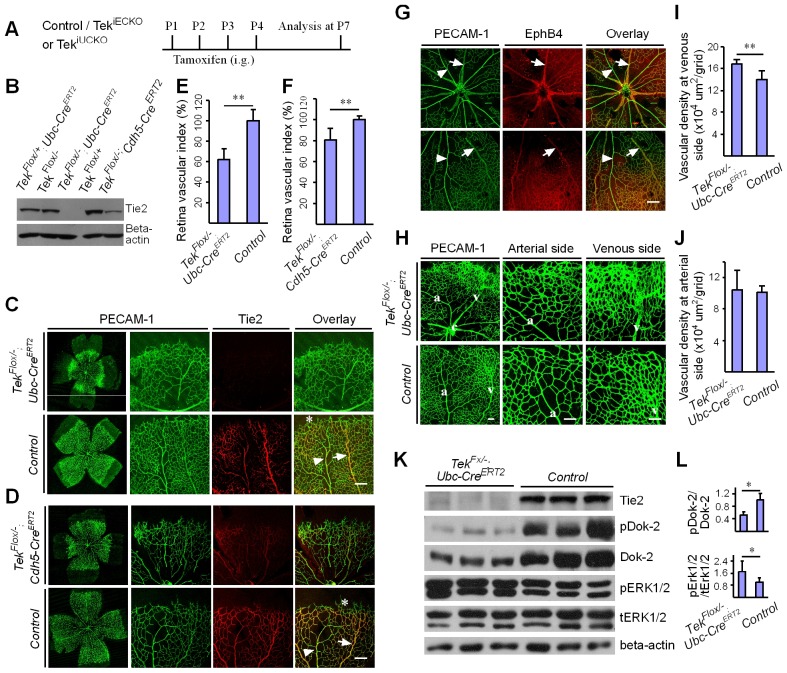
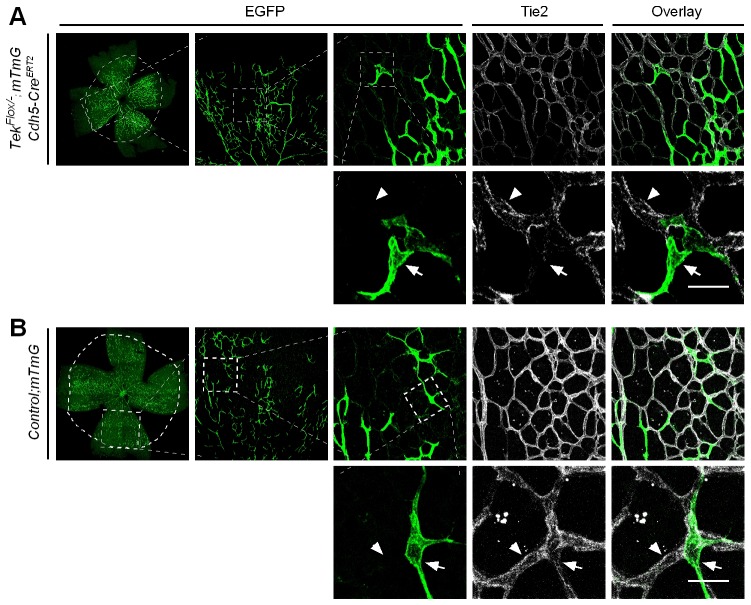
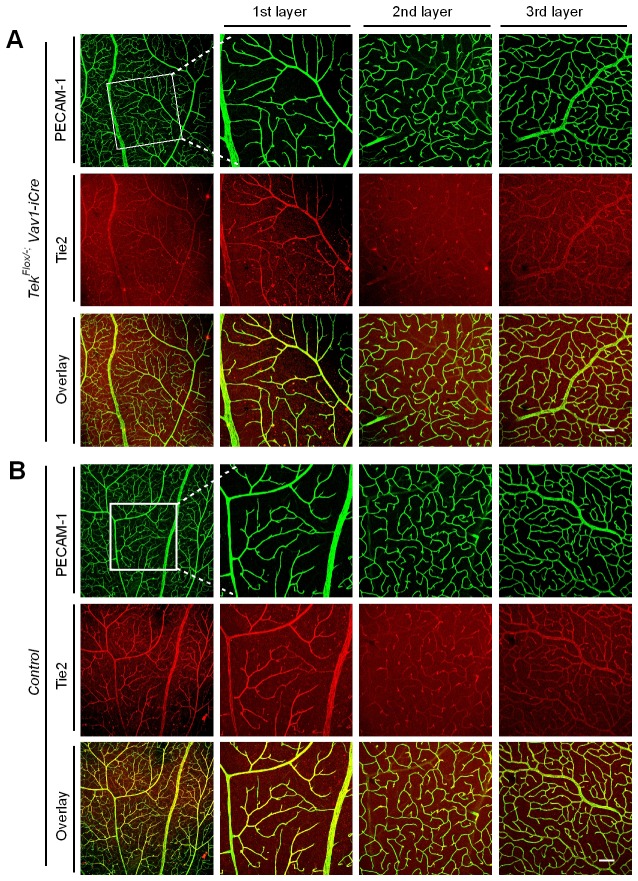
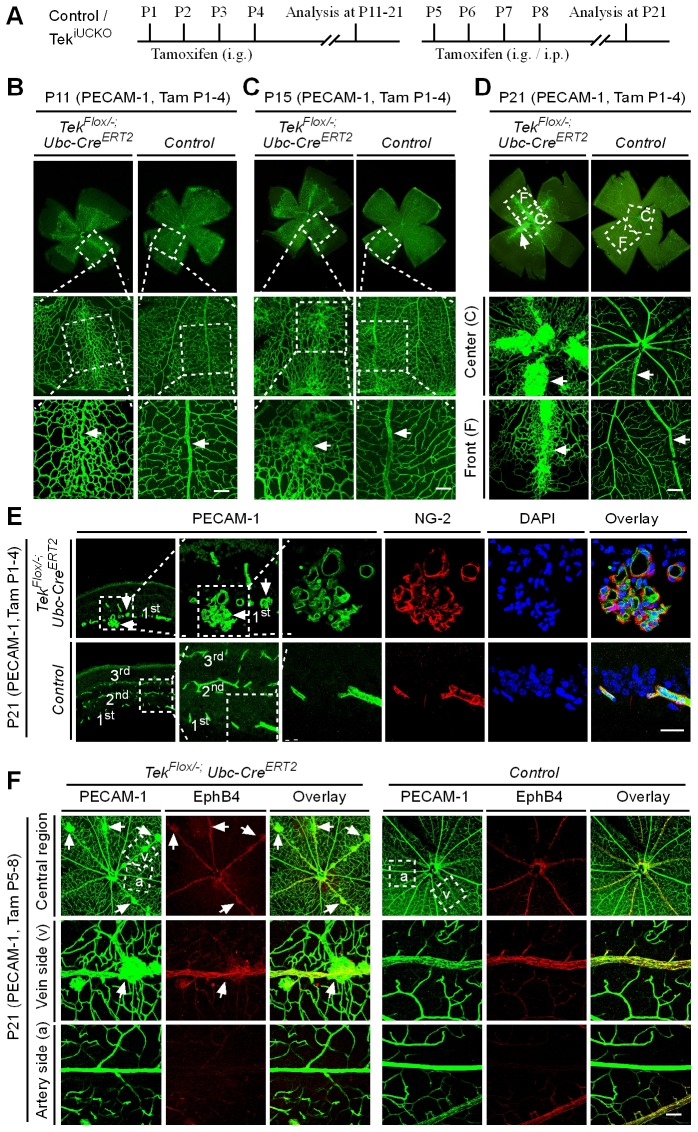
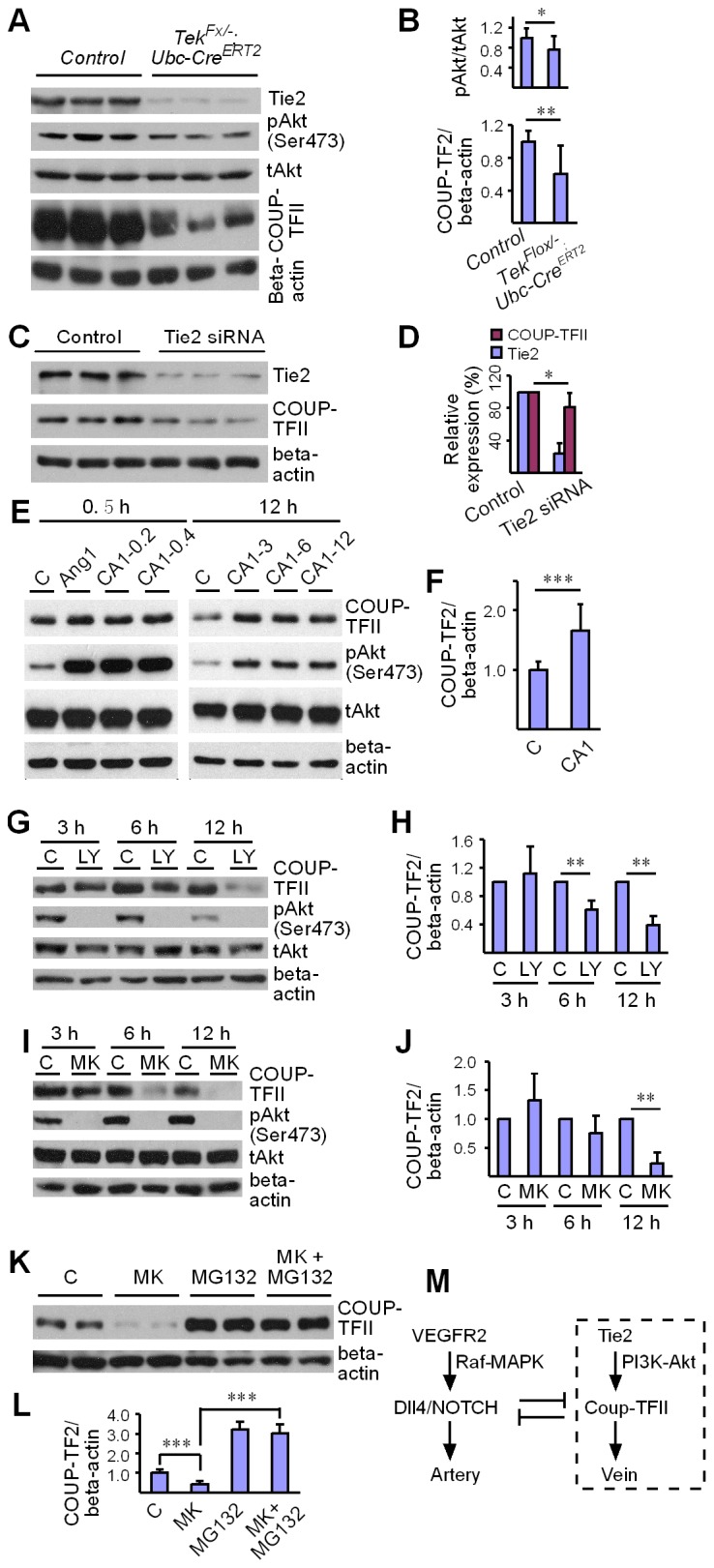
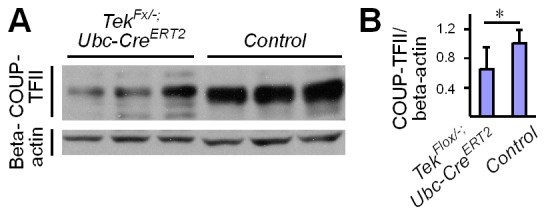
Mechanisms underlying the vein development remain largely unknown. Tie2 signaling mediates endothelial cell (EC) survival and vascular maturation and its activating mutations are linked to venous malformations. Here we show that vein formation are disrupted in mouse skin and mesentery when Tie2 signals are diminished by targeted deletion of Tek either ubiquitously or specifically in embryonic ECs. Postnatal Tie2 attenuation resulted in the degeneration of newly formed veins followed by the formation of haemangioma-like vascular tufts in retina and venous tortuosity. Mechanistically, Tie2 insufficiency compromised venous EC identity, as indicated by a significant decrease of COUP-TFII protein level, a key regulator in venogenesis. Consistently, angiopoietin-1 stimulation increased COUP-TFII in cultured ECs, while Tie2 knockdown or blockade of Tie2 downstream PI3K/Akt pathway reduced COUP-TFII which could be reverted by the proteasome inhibition. Together, our results imply that Tie2 is essential for venous specification and maintenance via Akt mediated stabilization of COUP-TFII.
Keywords: COUP-TFII; Tie2; developmental biology; mouse; stem cells; vein specification.
Conflict of interest statement
KA: Reviewing editor, eLife. The other authors declare that no competing interests exist.
Figures
Similar articles
-
Endothelial TIE1 Restricts Angiogenic Sprouting to Coordinate Vein Assembly in Synergy With Its Homologue TIE2.Arterioscler Thromb Vasc Biol. 2023 Aug;43(8):e323-e338. doi: 10.1161/ATVBAHA.122.318860. Epub 2023 Jun 15. Arterioscler Thromb Vasc Biol. 2023. PMID: 37317851
-
BRG1 promotes COUP-TFII expression and venous specification during embryonic vascular development.Development. 2013 Mar;140(6):1272-81. doi: 10.1242/dev.087379. Epub 2013 Feb 13. Development. 2013. PMID: 23406903 Free PMC article.
-
COUP-TFII regulates tumor growth and metastasis by modulating tumor angiogenesis.Proc Natl Acad Sci U S A. 2010 Feb 23;107(8):3687-92. doi: 10.1073/pnas.0914619107. Epub 2010 Feb 3. Proc Natl Acad Sci U S A. 2010. PMID: 20133706 Free PMC article.
-
Arterial-venous specification during development.Circ Res. 2009 Mar 13;104(5):576-88. doi: 10.1161/CIRCRESAHA.108.188805. Circ Res. 2009. PMID: 19286613 Review.
-
Molecular regulation of arteriovenous endothelial cell specification.F1000Res. 2019 Jul 29;8:F1000 Faculty Rev-1208. doi: 10.12688/f1000research.16701.1. eCollection 2019. F1000Res. 2019. PMID: 31448079 Free PMC article. Review.
Cited by
-
Vessel Enlargement in Development and Pathophysiology.Front Physiol. 2021 Feb 25;12:639645. doi: 10.3389/fphys.2021.639645. eCollection 2021. Front Physiol. 2021. PMID: 33716786 Free PMC article. Review.
-
Receptor tyrosine kinase inhibitors in cancer.Cell Mol Life Sci. 2023 Mar 22;80(4):104. doi: 10.1007/s00018-023-04729-4. Cell Mol Life Sci. 2023. PMID: 36947256 Free PMC article. Review.
-
Transcriptional regulators of arterial and venous identity in the developing mammalian embryo.Curr Opin Physiol. 2023 Oct;35:None. doi: 10.1016/j.cophys.2023.100691. Curr Opin Physiol. 2023. PMID: 38328689 Free PMC article. Review.
-
Endothelial Cell Development and Its Application to Regenerative Medicine.Circ Res. 2019 Aug 2;125(4):489-501. doi: 10.1161/CIRCRESAHA.119.311405. Epub 2019 Aug 1. Circ Res. 2019. PMID: 31518171 Free PMC article. Review.
-
Endothelial Heterogeneity in Development and Wound Healing.Cells. 2021 Sep 7;10(9):2338. doi: 10.3390/cells10092338. Cells. 2021. PMID: 34571987 Free PMC article. Review.
References
-
- Arita Y, Nakaoka Y, Matsunaga T, Kidoya H, Yamamizu K, Arima Y, Kataoka-Hashimoto T, Ikeoka K, Yasui T, Masaki T, Yamamoto K, Higuchi K, Park JS, Shirai M, Nishiyama K, Yamagishi H, Otsu K, Kurihara H, Minami T, Yamauchi-Takihara K, Koh GY, Mochizuki N, Takakura N, Sakata Y, Yamashita JK, Komuro I. Myocardium-derived angiopoietin-1 is essential for coronary vein formation in the developing heart. Nature Communications. 2014;5:4552. doi: 10.1038/ncomms5552. - DOI - PMC - PubMed
-
- Boscolo E, Limaye N, Huang L, Kang KT, Soblet J, Uebelhoer M, Mendola A, Natynki M, Seront E, Dupont S, Hammer J, Legrand C, Brugnara C, Eklund L, Vikkula M, Bischoff J, Boon LM. Rapamycin improves TIE2-mutated venous malformation in murine model and human subjects. Journal of Clinical Investigation. 2015;125:3491–3504. doi: 10.1172/JCI76004. - DOI - PMC - PubMed
-
- Corada M, Nyqvist D, Orsenigo F, Caprini A, Giampietro C, Taketo MM, Iruela-Arispe ML, Adams RH, Dejana E. The wnt/beta-catenin pathway modulates vascular remodeling and specification by upregulating Dll4/Notch signaling. Developmental Cell. 2010;18:938–949. doi: 10.1016/j.devcel.2010.05.006. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
